

Direct inhibition of CDK9 blocks HIV-1 replication without preventing T-cell activation in primary human peripheral blood lymphocytes
- PMID: 17949927
- PMCID: PMC2222555
- DOI: 10.1016/j.gene.2007.09.010
Direct inhibition of CDK9 blocks HIV-1 replication without preventing T-cell activation in primary human peripheral blood lymphocytes
Abstract
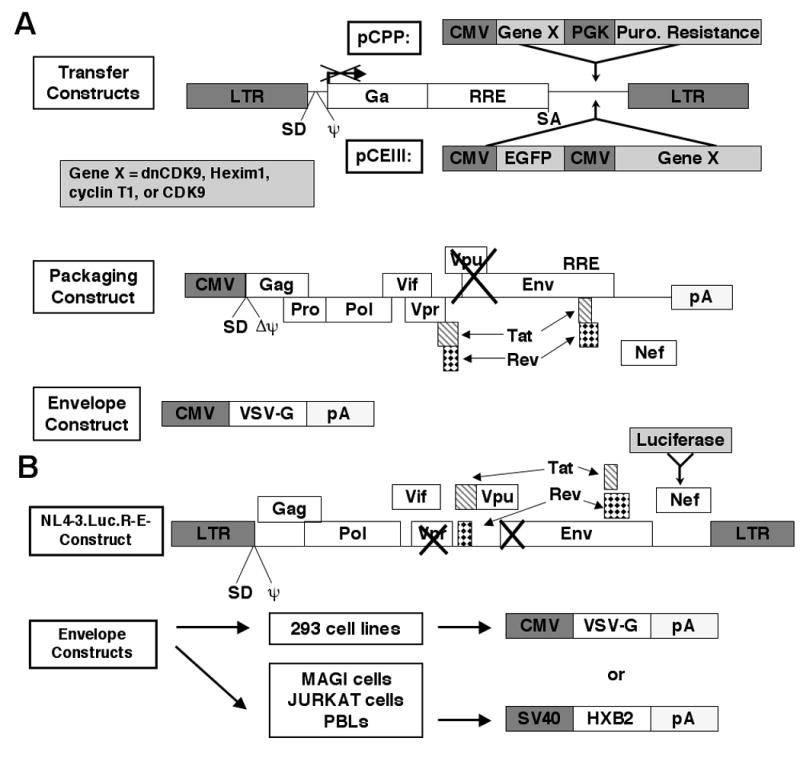
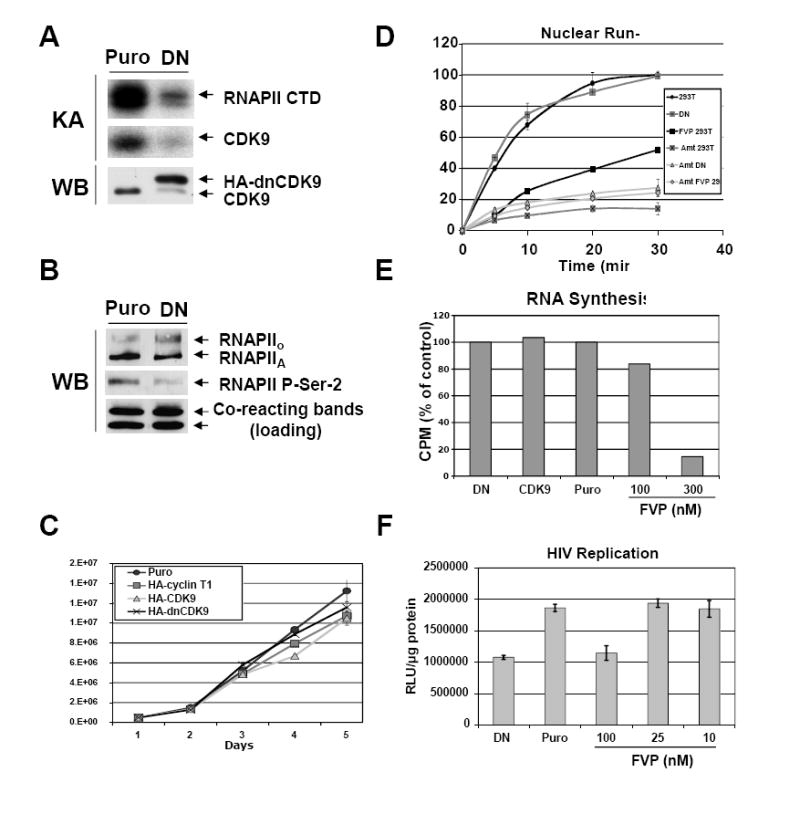
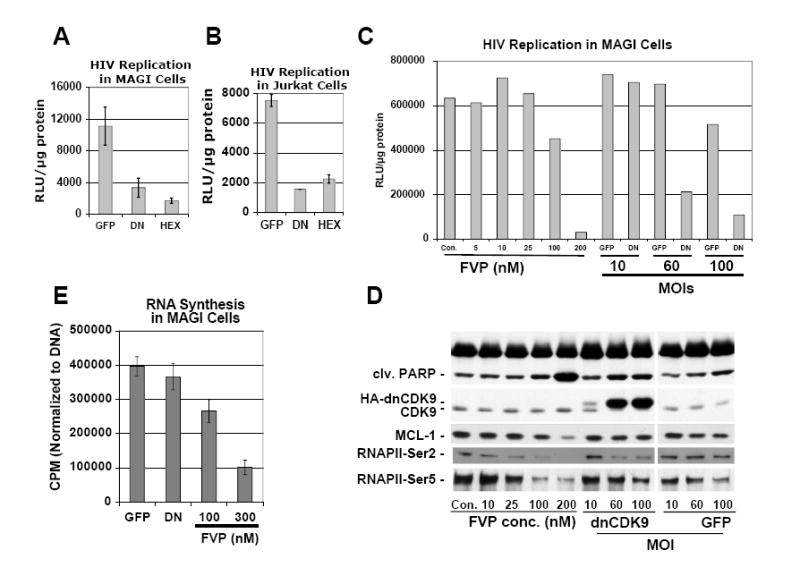
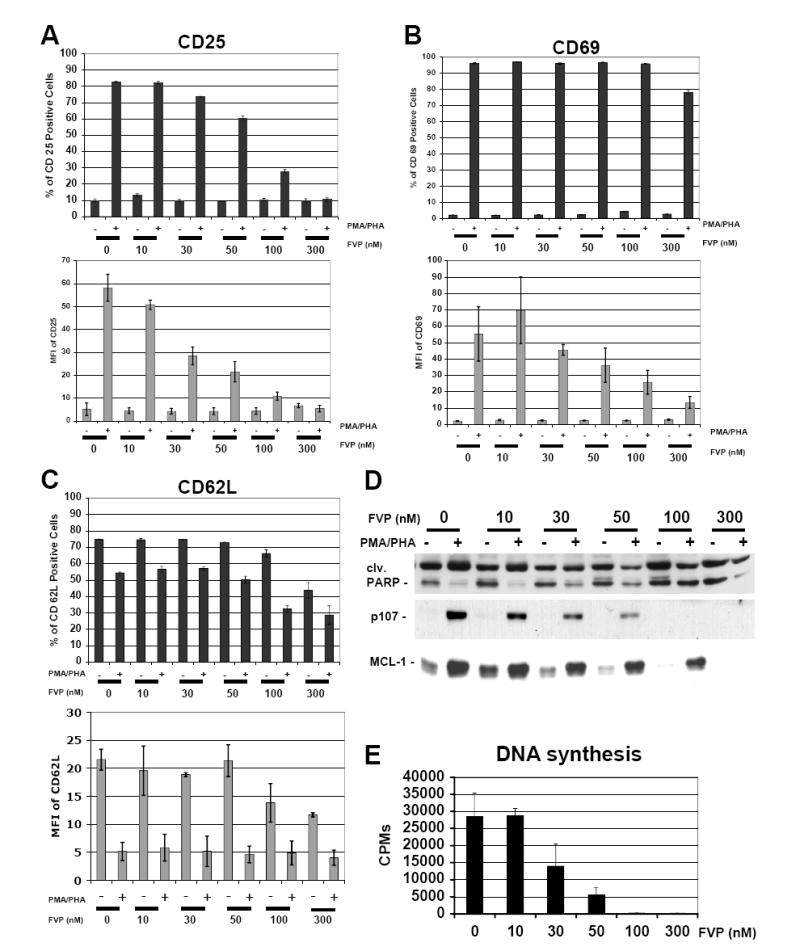
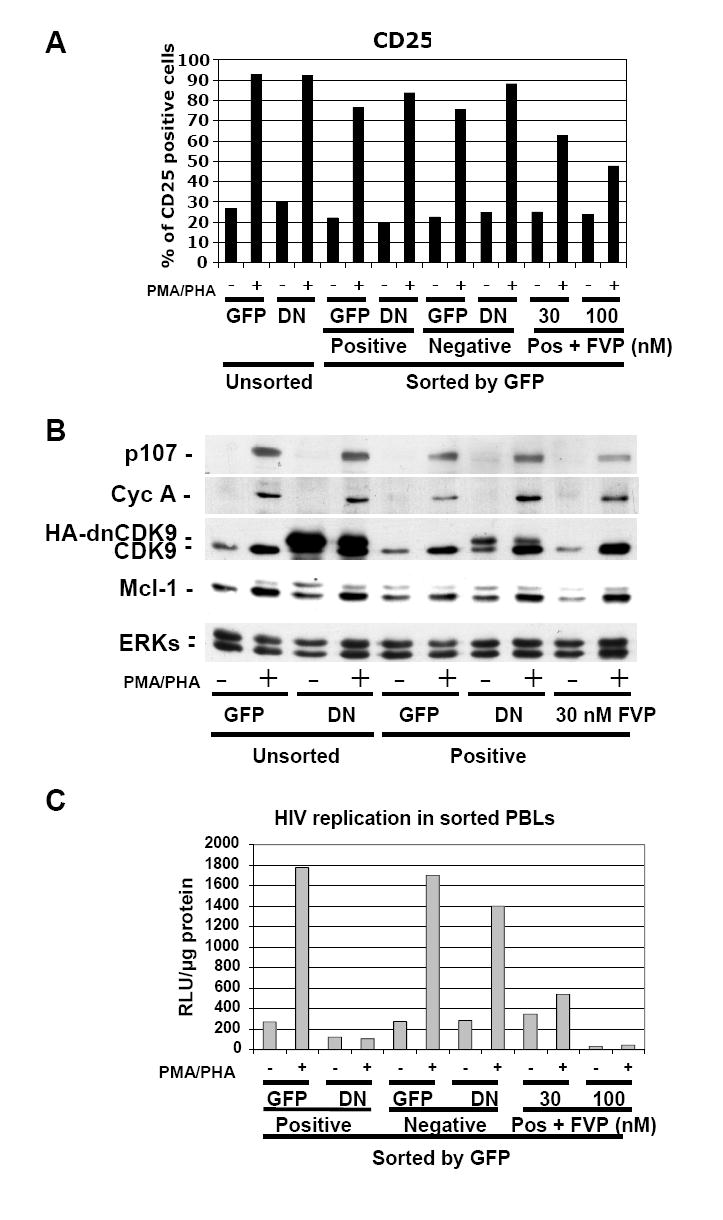
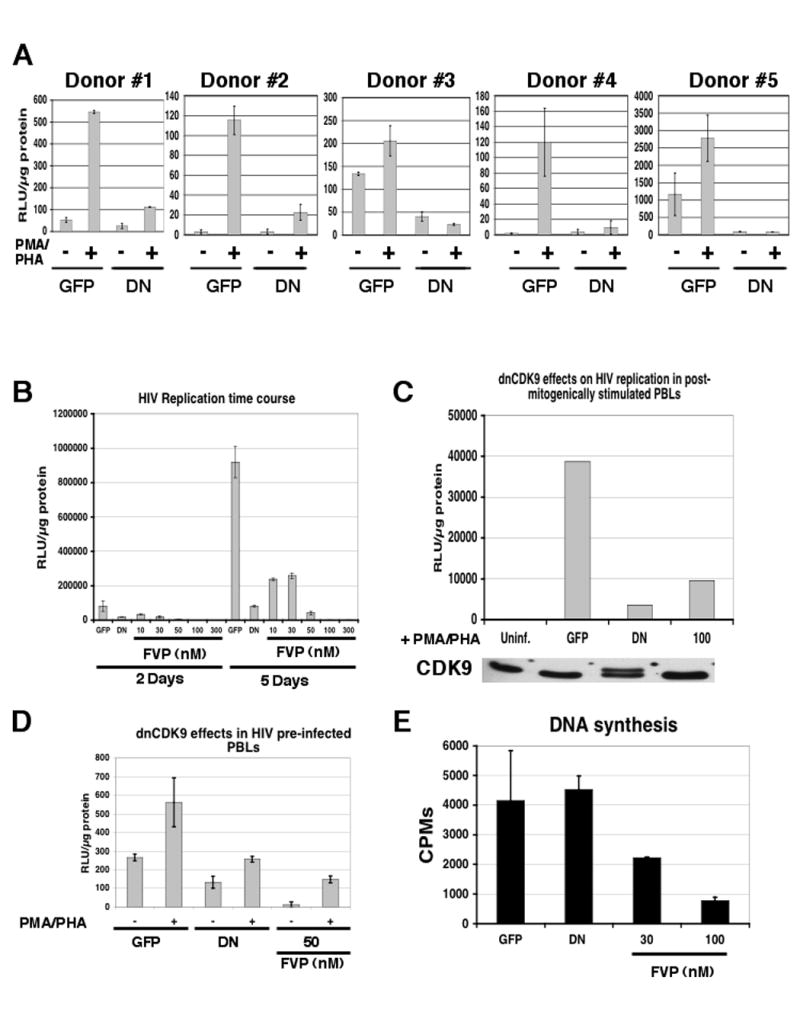
HIV-1 transcription is essential for the virus replication cycle. HIV-1 Tat is a viral transactivator that strongly stimulates the processivity of RNA polymerase II (RNAPII) via recruitment of the cyclin T1/CDK9 positive transcription elongation factor, which phosphorylates the C-terminal domain (CTD) of RNAPII. Consistently, HIV-1 replication in transformed cells is very sensitive to direct CDK9 inhibition. Thus, CDK9 could be a potential target for anti-HIV-1 therapy. A clearer understanding of the requirements for CDK9 activity in primary human T cells is needed to assess whether the CDK9-dependent step in HIV-1 transcription can be targeted clinically. We have investigated the effects of limiting CDK9 activity with recombinant lentiviruses expressing a dominant-negative form of CDK9 (HA-dnCDK9) in peripheral blood lymphocytes (PBLs) and other cells. Our results show that direct inhibition of CDK9 potently inhibits HIV-1 replication in single-round infection assays with little to undetectable effects on RNAPII transcription, RNA synthesis, proliferation and viability. In PBLs purified from multiple donors, direct inhibition of CDK9 activity blocks HIV-1 replication/transcription but does not prevent T-cell activation, as determined via measurement of cell surface and cell cycle entry and progression markers, and DNA synthesis. We have also compared the effects of HA-dnCDK9 to flavopiridol (FVP), a general CDK inhibitor that potently inhibits CDK9. In contrast to HA-dnCDK9, FVP interferes with key cellular processes at concentrations that inhibit HIV-1 replication with potency similar to HA-dnCDK9. In particular, FVP inhibits several T-cell activation markers and DNA synthesis in primary PBLs at the minimal concentrations required to inhibit HIV-1 replication. Our results imply that small pharmacological compounds targeting CDK9 with enhanced selectivity could be developed into effective anti-HIV-1 therapeutic drugs.
Figures
Similar articles
-
CDK9 inhibition strategy defines distinct sets of target genes.BMC Res Notes. 2014 May 16;7:301. doi: 10.1186/1756-0500-7-301. BMC Res Notes. 2014. PMID: 24886624 Free PMC article.
-
Selective control of gene expression by CDK9 in human cells.J Cell Physiol. 2010 Jan;222(1):200-8. doi: 10.1002/jcp.21938. J Cell Physiol. 2010. PMID: 19780058 Free PMC article.
-
Upregulation of cyclin T1/CDK9 complexes during T cell activation.Oncogene. 1998 Dec 17;17(24):3093-102. doi: 10.1038/sj.onc.1202548. Oncogene. 1998. PMID: 9872325
-
Mechanisms controlling CDK9 activity.Front Biosci. 2006 Sep 1;11:2598-613. doi: 10.2741/1994. Front Biosci. 2006. PMID: 16720337 Review.
-
CDK9 a potential target for drug development.Med Chem. 2008 May;4(3):210-8. doi: 10.2174/157340608784325205. Med Chem. 2008. PMID: 18473913 Review.
Cited by
-
P-TEFb goes viral.Inside Cell. 2016 Apr;1(2):106-116. doi: 10.1002/icl3.1037. Epub 2015 Nov 25. Inside Cell. 2016. PMID: 27398404 Free PMC article. Review.
-
Effects of Cyclin Dependent Kinase 9 inhibition on zebrafish larvae.Cell Cycle. 2016 Nov 16;15(22):3060-3069. doi: 10.1080/15384101.2016.1231283. Epub 2016 Oct 7. Cell Cycle. 2016. PMID: 27715402 Free PMC article.
-
Reduce and Control: A Combinatorial Strategy for Achieving Sustained HIV Remissions in the Absence of Antiretroviral Therapy.Viruses. 2020 Feb 8;12(2):188. doi: 10.3390/v12020188. Viruses. 2020. PMID: 32046251 Free PMC article. Review.
-
Inhibition of cdk9 during herpes simplex virus 1 infection impedes viral transcription.PLoS One. 2013 Oct 18;8(10):e79007. doi: 10.1371/journal.pone.0079007. eCollection 2013. PLoS One. 2013. PMID: 24205359 Free PMC article.
-
Latent HIV-1 infection occurs in multiple subsets of hematopoietic progenitor cells and is reversed by NF-κB activation.J Virol. 2012 Sep;86(17):9337-50. doi: 10.1128/JVI.00895-12. Epub 2012 Jun 20. J Virol. 2012. PMID: 22718820 Free PMC article.
References
-
- Blagosklonny MV. Flavopiridol, an inhibitor of transcription: implications, problems and solutions. Cell Cycle. 2004;3:1537–42. - PubMed
-
- Chao SH, Fujinaga K, Marion JE, Taube R, Sausville EA, Senderowicz AM, Peterlin BM, Price DH. Flavopiridol inhibits P-TEFb and blocks HIV-1 replication. J Biol Chem. 2000;275:28345–8. - PubMed
-
- Chao SH, Price DH. Flavopiridol inactivates P-TEFb and blocks most RNA polymerase II transcription in vivo. J Biol Chem. 2001;276:31793–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- M01 RR000349-350343/RR/NCRR NIH HHS/United States
- R01 AI045450-04/AI/NIAID NIH HHS/United States
- R01 CA078499-08/CA/NCI NIH HHS/United States
- M01 RR000349/RR/NCRR NIH HHS/United States
- R01 AI045450-03/AI/NIAID NIH HHS/United States
- R01 AI045450/AI/NIAID NIH HHS/United States
- K02 AI001823-01/AI/NIAID NIH HHS/United States
- K02 AI001823-04/AI/NIAID NIH HHS/United States
- K02 AI01823/AI/NIAID NIH HHS/United States
- R01 AI045450-02/AI/NIAID NIH HHS/United States
- R01 AI45450/AI/NIAID NIH HHS/United States
- R01 AI045450-01/AI/NIAID NIH HHS/United States
- K02 AI001823-02/AI/NIAID NIH HHS/United States
- M01 RR000349-330343/RR/NCRR NIH HHS/United States
- K02 AI001823/AI/NIAID NIH HHS/United States
- R01 CA78499/CA/NCI NIH HHS/United States
- K02 AI001823-05/AI/NIAID NIH HHS/United States
- K02 AI001823-03/AI/NIAID NIH HHS/United States
- M01 RR00349/RR/NCRR NIH HHS/United States
- R24 CA88261-01/CA/NCI NIH HHS/United States
- R01 CA078499/CA/NCI NIH HHS/United States
- M01 RR000349-340343/RR/NCRR NIH HHS/United States
- R24 CA088261/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
