

doi: 10.1038/nm1651.
Epub 2007 Sep 9.
Human TH17 lymphocytes promote blood-brain barrier disruption and central nervous system inflammation
Affiliations
- PMID: 17828272
- PMCID: PMC5114125
- DOI: 10.1038/nm1651
Item in Clipboard
Human TH17 lymphocytes promote blood-brain barrier disruption and central nervous system inflammation
Nat Med.
2007 Oct.
Abstract
T(H)17 lymphocytes appear to be essential in the pathogenesis of numerous inflammatory diseases. We demonstrate here the expression of IL-17 and IL-22 receptors on blood-brain barrier endothelial cells (BBB-ECs) in multiple sclerosis lesions, and show that IL-17 and IL-22 disrupt BBB tight junctions in vitro and in vivo. Furthermore, T(H)17 lymphocytes transmigrate efficiently across BBB-ECs, highly express granzyme B, kill human neurons and promote central nervous system inflammation through CD4+ lymphocyte recruitment.
Figures
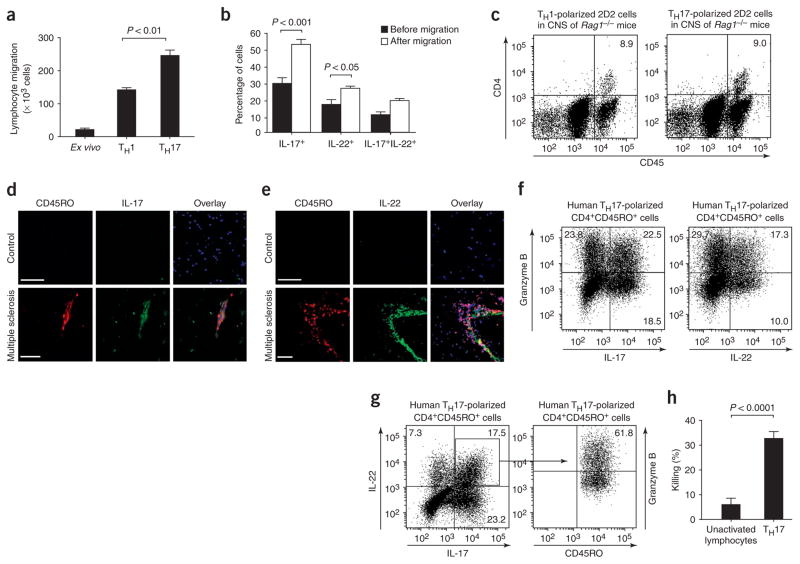
TH17 lymphocytes migrate efficiently across the BBB in vitro and in vivo and kill human neurons. (a) Human CD4+CD45RO+ TH17, CD4+ TH1 (both generated in vitro, see Supplementary Methods) and ex vivo CD4+ lymphocytes were allowed to migrate across human BBB-ECs in a modified Boyden chamber assay (ref. and Supplementary Methods) for 18 h. Significantly more TH17 lymphocytes migrated than either TH1 or ex vivo CD4+ lymphocytes. (b) CD4+CD45RO+ TH17 lymphocytes were allowed to migrate across human BBB-ECs for 18 h. Cells were stained for IL-17 and IL-22 before and after migration. The cytokine profile revealed the preferential migration of IL-17+ and IL-22+ lymphocytes. (c) Immune cells from lymph nodes and spleen of MOG35–55-immunized 2D2 mice were polarized toward TH1 or TH17 and transferred to Rag1−/− mice, and CD45hiCD4+ lymphocytes were isolated from the CNS 7 d after transfer. Shown is a representative flow cytometry dot plot of CNS cell content from Rag1−/− mice injected with either TH1- (left) or TH17-polarized (right) 2D2 lymphocytes (n = 4 mice per group). (d) Human CNS postmortem material from unaffected individuals (control, non-inflamed; above) and heavily infiltrated CNS material from individuals with multiple sclerosis (below) were immunostained for CD45RO (red), IL-17 (green) and nuclear stain TO-PRO3 (blue). Confocal microscopy imaging confirmed the presence of IL-17+CD45RO+ cells (yellow) in infiltrated multiple sclerosis lesions but not in control CNS. Bar, 75 μm. (e) Similarly, IL-22+CD45RO+ staining was observed in multiple sclerosis lesions, but not in control CNS material. (f) Human CD4+CD45RO+ TH17–polarized lymphocytes were stained for CD45RO, IL-17, IL-22 and granzyme B. Both IL-17– and IL-22–producing lymphocytes expressed granzyme B (22.5% and 17.3%, respectively). (g) More than 60% of IL-17+IL-22+ lymphocytes highly expressed granzyme B. Granzyme A and perforin were not detected in TH17 cells, whether or not these cells produced IL-22 (data not shown). (h) The cytotoxic activity of TH17 lymphocytes was assessed using neuron-enriched cultures obtained from human fetal CNS material and compared to that of unactivated T lymphocytes. All data shown are representative of the mean ± s.e.m. of three independent experiments.
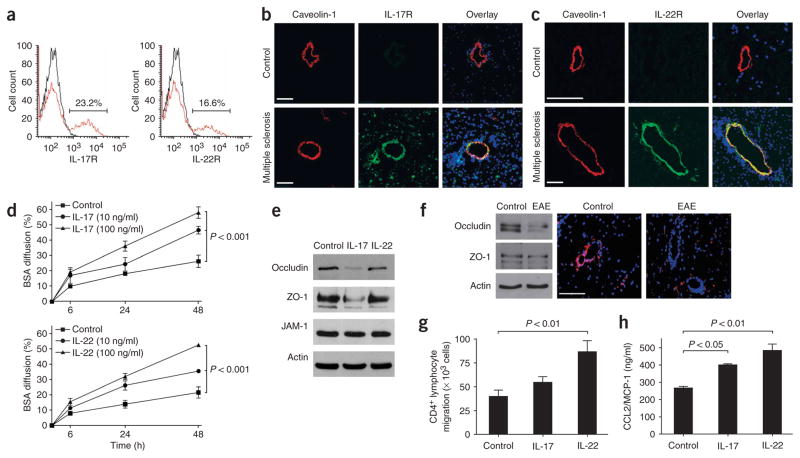
IL-17 and IL-22 receptors are expressed on human brain endothelium, and their activation permeabilizes the BBB. (a) Unactivated human BBB-ECs grown in primary culture were stained for IL-17R and IL-22R, revealing their expression on the surface of 23.2% and 16.6% of BBB-ECs, respectively. (b) Human CNS postmortem material from unaffected individuals (control, non-inflamed; above) and heavily infiltrated CNS material from individuals with multiple sclerosis (below) were immunostained for IL-17R (green), caveolin-1 (red) and nuclear stain TO-PRO3 (blue). Confocal microscopy imaging confirmed the expression of IL-17R on caveolin-1+ endothelium in inflamed CNS material. IL-17R expression was undetectable in control CNS material. Bar, 75 μm. (c) Similarly, IL-22R staining was observed on endothelial cells in multiple sclerosis lesions, but not in controls. (d) Human BBB-ECs were grown in Boyden chambers and treated with IL-17 (top) or IL-22 (bottom). Permeability of the monolayers was monitored with fluorescent BSA, showing that BBB-EC monolayer permeability increased after treatment with either IL-17 or IL-22. (e) Western blot for the tight-junction proteins occludin, ZO-1 and junction adhesion molecule (JAM)-1 from human BBB-ECs revealed disruption of occludin and ZO-1 by IL-17 (100 ng/ml, 18 h). (f) Western blot for tight-junction proteins in spinal cord homogenates of MOG35–55-immunized EAE mice revealed a similar reduction in occludin and ZO-1. In situ immunostaining for ZO-1 (red) and nuclear stain TO-PRO3 (blue) in normal-appearing cerebellar white matter (control) and in infiltrated and demyelinated cerebellar lesions from C57BL/6 mice immunized with MOG35–55 (EAE, grade 4). Confocal microscopy imaging confirmed disruption of ZO-1 around infiltrated vessels. Bar, 75 μm. (g) Freshly isolated peripheral blood human CD4+ lymphocytes were allowed to migrate for 18 h across IL-17– (100 ng/ml) or IL-22–treated (100 ng/ml) human BBB-ECs. Both cytokines promoted migration of human ex vivo CD4+ lymphocytes across human BBB-ECs, as compared to control. (h) CCL2 (or MCP-1) secretion by human BBB-ECs was assessed by ELISA in untreated and IL-17– or IL-22–treated cultures (100 ng/ml, 18 h). Both IL-17 and IL-22 upregulate CCL2 secretion by human BBB-ECs. All data shown represent the mean ± s.e.m. from three independent experiments performed in triplicate.
Similar articles
-
Statins reduce human blood-brain barrier permeability and restrict leukocyte migration: relevance to multiple sclerosis.Ann Neurol. 2006 Jul;60(1):45-55. doi: 10.1002/ana.20875. Ann Neurol. 2006. PMID: 16729291
-
Cellular mechanisms of IL-17-induced blood-brain barrier disruption.FASEB J. 2010 Apr;24(4):1023-34. doi: 10.1096/fj.09-141978. Epub 2009 Nov 25. FASEB J. 2010. PMID: 19940258
-
Preferential recruitment of interferon-gamma-expressing TH17 cells in multiple sclerosis.Ann Neurol. 2009 Sep;66(3):390-402. doi: 10.1002/ana.21748. Ann Neurol. 2009. PMID: 19810097
-
The blood-brain-barrier in multiple sclerosis: functional roles and therapeutic targeting.Autoimmunity. 2007 Mar;40(2):148-60. doi: 10.1080/08916930601183522. Autoimmunity. 2007. PMID: 17453713 Review.
-
Molecular and cellular permeability control at the blood-brain barrier.Brain Res Brain Res Rev. 2001 Oct;36(2-3):258-64. doi: 10.1016/s0165-0173(01)00102-3. Brain Res Brain Res Rev. 2001. PMID: 11690623 Review.
Cited by
-
Integrated omics profiling reveals systemic dysregulation and potential biomarkers in the blood of patients with neuromyelitis optica spectrum disorders.J Transl Med. 2024 Nov 1;22(1):989. doi: 10.1186/s12967-024-05801-8. J Transl Med. 2024. PMID: 39487546 Free PMC article.
-
Expansion in CD39⁺ CD4⁺ immunoregulatory t cells and rarity of Th17 cells in HTLV-1 infected patients is associated with neurological complications.PLoS Negl Trop Dis. 2013;7(2):e2028. doi: 10.1371/journal.pntd.0002028. Epub 2013 Feb 7. PLoS Negl Trop Dis. 2013. PMID: 23409198 Free PMC article.
-
Pericytes regulate vascular immune homeostasis in the CNS.Proc Natl Acad Sci U S A. 2021 Mar 9;118(10):e2016587118. doi: 10.1073/pnas.2016587118. Proc Natl Acad Sci U S A. 2021. PMID: 33653955 Free PMC article.
-
Current Immunological and Clinical Perspective on Vaccinations in Multiple Sclerosis Patients: Are They Safe after All?Int J Mol Sci. 2021 Apr 8;22(8):3859. doi: 10.3390/ijms22083859. Int J Mol Sci. 2021. PMID: 33917860 Free PMC article. Review.
-
Fine mapping and functional analysis of the multiple sclerosis risk gene CD6.PLoS One. 2013 Apr 24;8(4):e62376. doi: 10.1371/journal.pone.0062376. Print 2013. PLoS One. 2013. PMID: 23638056 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
