

The spindle pole bodies facilitate nuclear envelope division during closed mitosis in fission yeast
- PMID: 17579515
- PMCID: PMC1892572
- DOI: 10.1371/journal.pbio.0050170
The spindle pole bodies facilitate nuclear envelope division during closed mitosis in fission yeast
Abstract
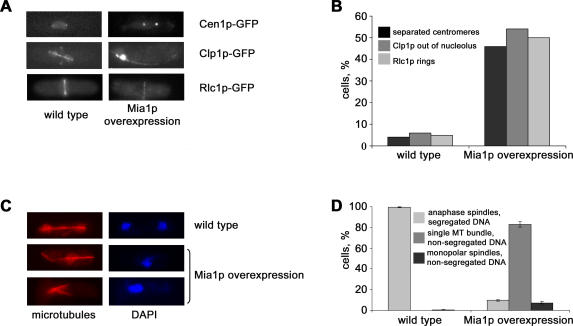
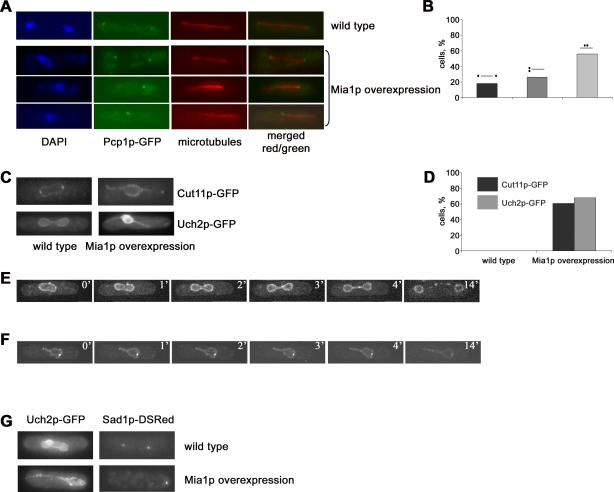
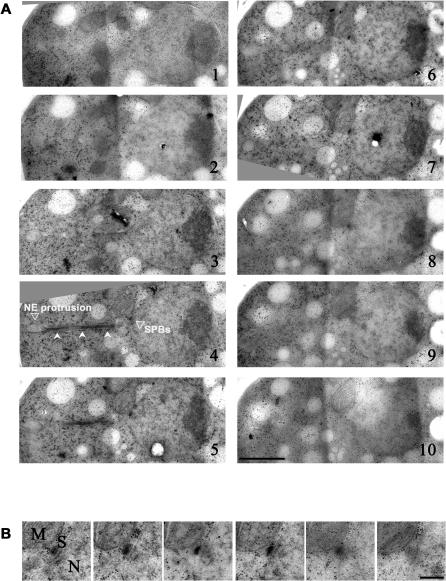
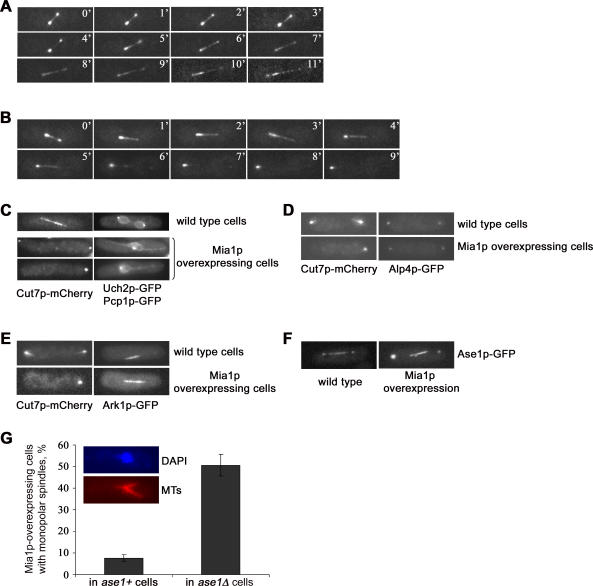
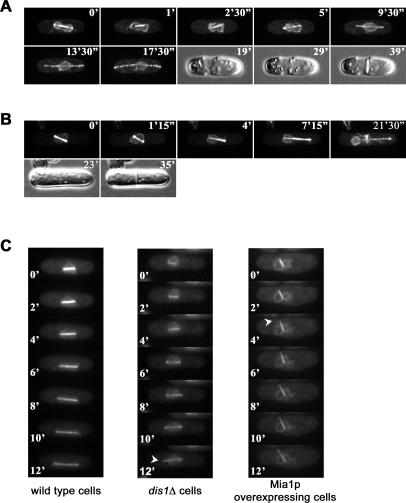
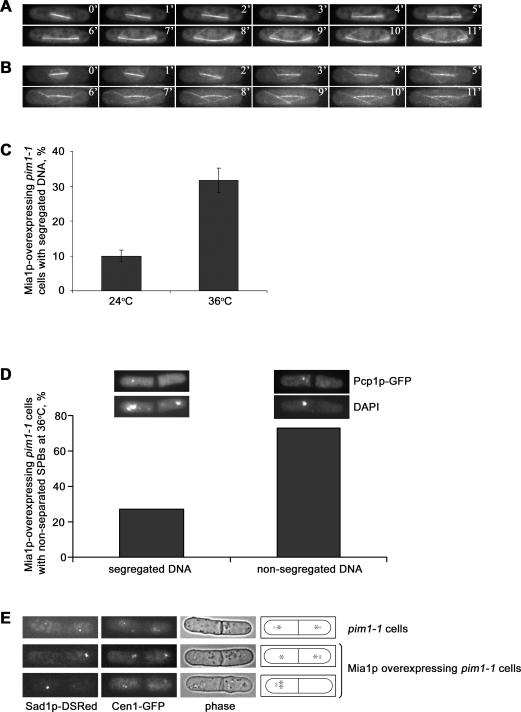
Many organisms divide chromosomes within the confines of the nuclear envelope (NE) in a process known as closed mitosis. Thus, they must ensure coordination between segregation of the genetic material and division of the NE itself. Although many years of work have led to a reasonably clear understanding of mitotic spindle function in chromosome segregation, the NE division mechanism remains obscure. Here, we show that fission yeast cells overexpressing the transforming acid coiled coil (TACC)-related protein, Mia1p/Alp7p, failed to separate the spindle pole bodies (SPBs) at the onset of mitosis, but could assemble acentrosomal bipolar and antiparallel spindle structures. Most of these cells arrested in anaphase with fully extended spindles and nonsegregated chromosomes. Spindle poles that lacked the SPBs did not lead the division of the NE during spindle elongation, but deformed it, trapping the chromosomes within. When the SPBs were severed by laser microsurgery in wild-type cells, we observed analogous deformations of the NE by elongating spindle remnants, resulting in NE division failure. Analysis of dis1Delta cells that elongate spindles despite unattached kinetochores indicated that the SPBs were required for maintaining nuclear shape at anaphase onset. Strikingly, when the NE was disassembled by utilizing a temperature-sensitive allele of the Ran GEF, Pim1p, the abnormal spindles induced by Mia1p overexpression were capable of segregating sister chromatids to daughter cells, suggesting that the failure to divide the NE prevents chromosome partitioning. Our results imply that the SPBs preclude deformation of the NE during spindle elongation and thus serve as specialized structures enabling nuclear division during closed mitosis in fission yeast.
Conflict of interest statement
Figures
Similar articles
-
cut11(+): A gene required for cell cycle-dependent spindle pole body anchoring in the nuclear envelope and bipolar spindle formation in Schizosaccharomyces pombe.Mol Biol Cell. 1998 Oct;9(10):2839-55. doi: 10.1091/mbc.9.10.2839. Mol Biol Cell. 1998. PMID: 9763447 Free PMC article.
-
Fission yeast cells undergo nuclear division in the absence of spindle microtubules.PLoS Biol. 2010 Oct 12;8(10):e1000512. doi: 10.1371/journal.pbio.1000512. PLoS Biol. 2010. PMID: 20967237 Free PMC article.
-
Nuclear shape, growth and integrity in the closed mitosis of fission yeast depend on the Ran-GTPase system, the spindle pole body and the endoplasmic reticulum.J Cell Sci. 2009 Jul 15;122(Pt 14):2464-72. doi: 10.1242/jcs.049999. J Cell Sci. 2009. PMID: 19571115 Free PMC article.
-
Big Lessons from Little Yeast: Budding and Fission Yeast Centrosome Structure, Duplication, and Function.Annu Rev Genet. 2017 Nov 27;51:361-383. doi: 10.1146/annurev-genet-120116-024733. Epub 2017 Sep 15. Annu Rev Genet. 2017. PMID: 28934593 Review.
-
Remodeling the nuclear membrane during closed mitosis.Curr Opin Cell Biol. 2013 Feb;25(1):142-8. doi: 10.1016/j.ceb.2012.09.001. Epub 2012 Oct 5. Curr Opin Cell Biol. 2013. PMID: 23040820 Review.
Cited by
-
Temporal Regulation of Lipin Activity Diverged to Account for Differences in Mitotic Programs.Curr Biol. 2016 Jan 25;26(2):237-243. doi: 10.1016/j.cub.2015.11.061. Epub 2016 Jan 7. Curr Biol. 2016. PMID: 26774782 Free PMC article.
-
Specific detection of fission yeast primary septum reveals septum and cleavage furrow ingression during early anaphase independent of mitosis completion.PLoS Genet. 2018 May 29;14(5):e1007388. doi: 10.1371/journal.pgen.1007388. eCollection 2018 May. PLoS Genet. 2018. PMID: 29813053 Free PMC article.
-
Nucleocytoplasmic transport in the midzone membrane domain controls yeast mitotic spindle disassembly.J Cell Biol. 2015 May 11;209(3):387-402. doi: 10.1083/jcb.201412144. J Cell Biol. 2015. PMID: 25963819 Free PMC article.
-
Mixing and matching nuclear envelope remodeling and spindle assembly strategies in the evolution of mitosis.Curr Opin Cell Biol. 2016 Aug;41:43-50. doi: 10.1016/j.ceb.2016.03.016. Epub 2016 Apr 7. Curr Opin Cell Biol. 2016. PMID: 27062548 Free PMC article. Review.
-
Vesicle-like biomechanics governs important aspects of nuclear geometry in fission yeast.PLoS One. 2007 Sep 26;2(9):e948. doi: 10.1371/journal.pone.0000948. PLoS One. 2007. PMID: 17895989 Free PMC article.
References
-
- McCully EK, Robinow CF. Mitosis in the fission yeast Schizosaccharomyces pombe: A comparative study with light and electron microscopy. J Cell Sci. 1971;9:475–507. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
