

Structure and analysis of a complex between SUMO and Ubc9 illustrates features of a conserved E2-Ubl interaction
- PMID: 17466333
- PMCID: PMC1940065
- DOI: 10.1016/j.jmb.2007.04.006
Structure and analysis of a complex between SUMO and Ubc9 illustrates features of a conserved E2-Ubl interaction
Abstract
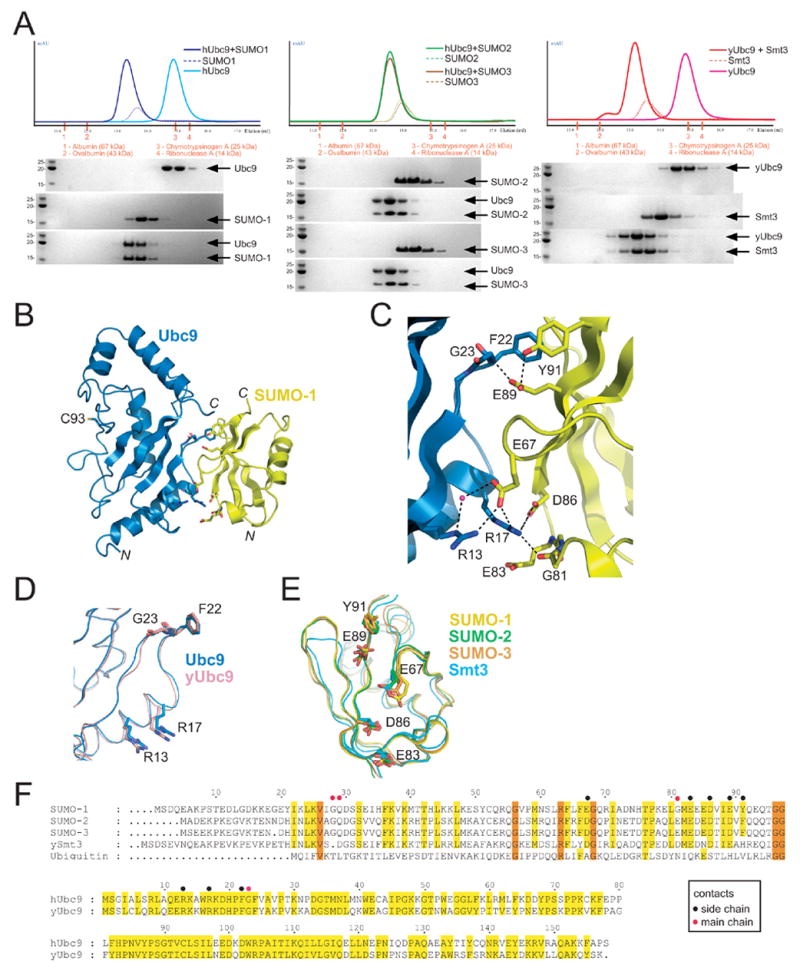
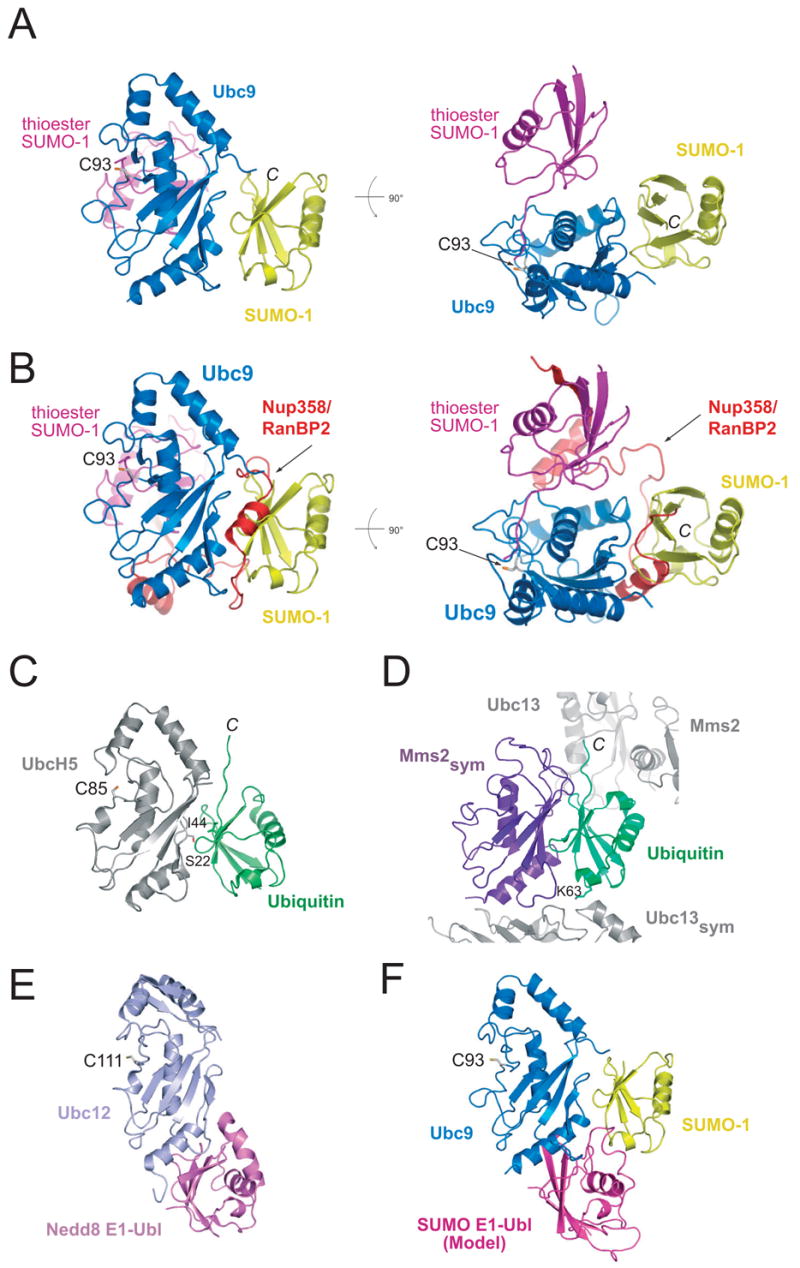
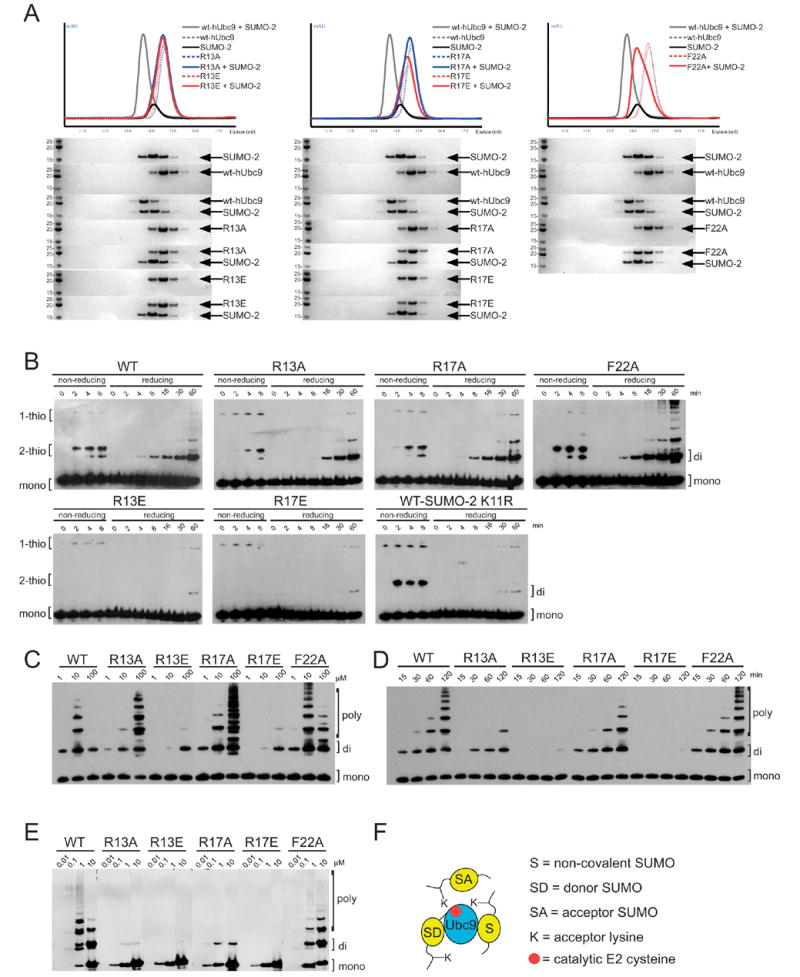
The SUMO E2 Ubc9 serves as a lynchpin in the SUMO conjugation pathway, interacting with the SUMO E1 during activation, with thioester linked SUMO after E1 transfer and with the substrate and SUMO E3 ligases during conjugation. Here, we describe the structure determination of a non-covalent complex between human Ubc9 and SUMO-1 at 2.4 A resolution. Non-covalent interactions between Ubc9 and SUMO are conserved in human and yeast insomuch as human Ubc9 interacts with each of the human SUMO isoforms, and yeast Ubc9 interacts with Smt3, the yeast SUMO ortholog. Structural comparisons reveal similarities to several other non-covalent complexes in the ubiquitin pathway, suggesting that the non-covalent Ubc9-SUMO interface may be important for poly-SUMO chain formation, for E2 recruitment to SUMO conjugated substrates, or for mediating E2 interactions with either E1 or E3 ligases. Biochemical analysis suggests that this surface is less important for E1 activation or di-SUMO-2 formation, but more important for E3 interactions and for poly-SUMO chain formation when the chain exceeds more than two SUMO proteins.
Figures
Similar articles
-
Crystal structure of UBA2(ufd)-Ubc9: insights into E1-E2 interactions in Sumo pathways.PLoS One. 2010 Dec 30;5(12):e15805. doi: 10.1371/journal.pone.0015805. PLoS One. 2010. PMID: 21209884 Free PMC article.
-
Insights into E3 ligase activity revealed by a SUMO-RanGAP1-Ubc9-Nup358 complex.Nature. 2005 Jun 2;435(7042):687-92. doi: 10.1038/nature03588. Nature. 2005. PMID: 15931224 Free PMC article.
-
Role of an N-terminal site of Ubc9 in SUMO-1, -2, and -3 binding and conjugation.Biochemistry. 2003 Aug 26;42(33):9959-69. doi: 10.1021/bi0345283. Biochemistry. 2003. PMID: 12924945
-
Protein interactions in the sumoylation cascade: lessons from X-ray structures.FEBS J. 2008 Jun;275(12):3003-15. doi: 10.1111/j.1742-4658.2008.06459.x. Epub 2008 May 17. FEBS J. 2008. PMID: 18492068 Review.
-
SUMO conjugation and cardiovascular development.Front Biosci (Landmark Ed). 2009 Jan 1;14(4):1219-29. doi: 10.2741/3304. Front Biosci (Landmark Ed). 2009. PMID: 19273126 Review.
Cited by
-
Identification of a new small ubiquitin-like modifier (SUMO)-interacting motif in the E3 ligase PIASy.J Biol Chem. 2017 Jun 16;292(24):10230-10238. doi: 10.1074/jbc.M117.789982. Epub 2017 Apr 28. J Biol Chem. 2017. PMID: 28455449 Free PMC article.
-
A new vertebrate SUMO enzyme family reveals insights into SUMO-chain assembly.Nat Struct Mol Biol. 2015 Dec;22(12):959-67. doi: 10.1038/nsmb.3114. Epub 2015 Nov 2. Nat Struct Mol Biol. 2015. PMID: 26524493
-
Site-specific inhibition of the small ubiquitin-like modifier (SUMO)-conjugating enzyme Ubc9 selectively impairs SUMO chain formation.J Biol Chem. 2017 Sep 15;292(37):15340-15351. doi: 10.1074/jbc.M117.794255. Epub 2017 Aug 7. J Biol Chem. 2017. PMID: 28784659 Free PMC article.
-
Crystal structure of UBA2(ufd)-Ubc9: insights into E1-E2 interactions in Sumo pathways.PLoS One. 2010 Dec 30;5(12):e15805. doi: 10.1371/journal.pone.0015805. PLoS One. 2010. PMID: 21209884 Free PMC article.
-
A novel SUMO1-specific interacting motif in dipeptidyl peptidase 9 (DPP9) that is important for enzymatic regulation.J Biol Chem. 2012 Dec 28;287(53):44320-9. doi: 10.1074/jbc.M112.397224. Epub 2012 Nov 14. J Biol Chem. 2012. PMID: 23152501 Free PMC article.
References
-
- Kerscher O, Felberbaum R, Hochstrasser M. Modification of proteins by ubiquitin and ubiquitin-like proteins. Annu Rev Cell Dev Biol. 2006;22:159–80. - PubMed
-
- Hershko A, Ciechanover A. The ubiquitin system. Annu Rev Biochem. 1998;67:425–79. - PubMed
-
- Hochstrasser M. There’s the rub: a novel ubiquitin-like modification linked to cell cycle regulation. Genes Dev. 1998;12:901–7. - PubMed
-
- Laney JD, Hochstrasser M. Substrate targeting in the ubiquitin system. Cell. 1999;97:427–30. - PubMed
-
- Schwartz DC, Hochstrasser M. A superfamily of protein tags: ubiquitin, SUMO and related modifiers. Trends Biochem Sci. 2003;28:321–8. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
