

Cross talk between notch and growth factor/cytokine signaling pathways in neural stem cells
- PMID: 17371842
- PMCID: PMC1900030
- DOI: 10.1128/MCB.00170-07
Cross talk between notch and growth factor/cytokine signaling pathways in neural stem cells
Abstract
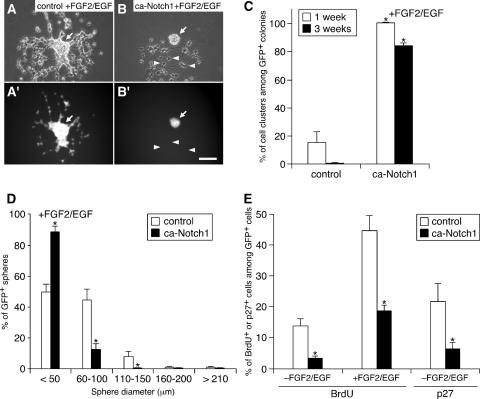
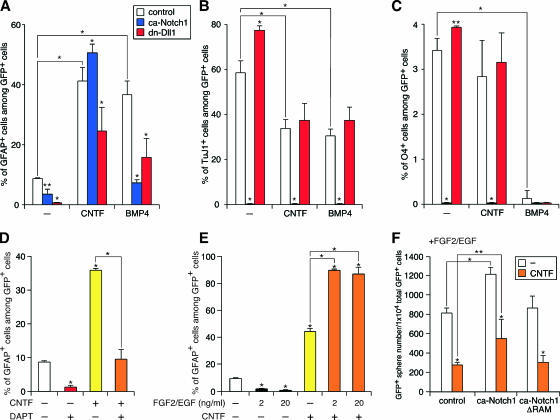
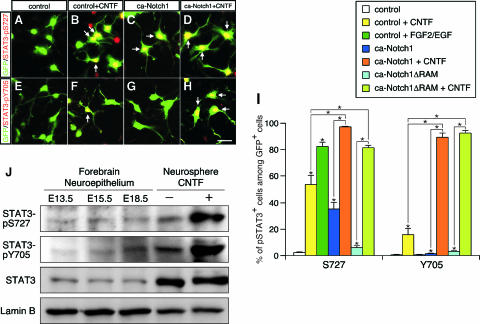
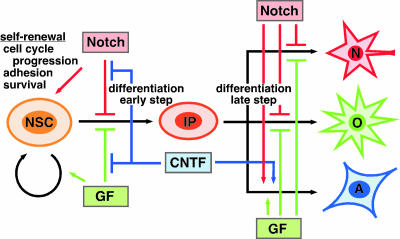
Precise control of proliferation and differentiation of multipotent neural stem cells (NSCs) is crucial for proper development of the nervous system. Although signaling through the cell surface receptor Notch has been implicated in many aspects of neural development, its role in NSCs remains elusive. Here we examined how the Notch pathway cross talks with signaling for growth factors and cytokines in controlling the self-renewal and differentiation of NSCs. Both Notch and growth factors were required for active proliferation of NSCs, but each of these signals was sufficient and independent of the other to inhibit differentiation of neurons and glia. Moreover, Notch signals could support the clonal self-renewing growth of NSCs in the absence of growth factors. This growth factor-independent action of Notch involved the regulation of the cell cycle and cell-cell interactions. During differentiation of NSCs, Notch signals promoted the generation of astrocytes in collaboration with ciliary neurotrophic factor and growth factors. Their cooperative actions were likely through synergistic phosphorylation of signal transducer and activator of transcription 3 on tyrosine at position 705 and serine at position 727. Our data suggest that distinct intracellular signaling pathways operate downstream of Notch for the self-renewal of NSCs and stimulation of astrogenesis.
Figures

Similar articles
-
Neurogenesis in the dentate gyrus depends on ciliary neurotrophic factor and signal transducer and activator of transcription 3 signaling.Stem Cells. 2009 Feb;27(2):431-41. doi: 10.1634/stemcells.2008-0234. Stem Cells. 2009. PMID: 19023034
-
Ciliary neurotrophic factor-mediated signaling regulates neuronal versus glial differentiation of retinal stem cells/progenitors by concentration-dependent recruitment of mitogen-activated protein kinase and Janus kinase-signal transducer and activator of transcription pathways in conjunction with Notch signaling.Stem Cells. 2008 Oct;26(10):2611-24. doi: 10.1634/stemcells.2008-0222. Epub 2008 Jul 31. Stem Cells. 2008. PMID: 18669911
-
Transcription factor RBP-J-mediated signaling represses the differentiation of neural stem cells into intermediate neural progenitors.Mol Cell Neurosci. 2009 Apr;40(4):442-50. doi: 10.1016/j.mcn.2008.12.008. Epub 2009 Jan 6. Mol Cell Neurosci. 2009. PMID: 19168137
-
Transcriptional regulatory networks associated with self-renewal and differentiation of neural stem cells.J Cell Physiol. 2010 Nov;225(2):337-47. doi: 10.1002/jcp.22294. J Cell Physiol. 2010. PMID: 20607797 Review.
-
[Research progress of neural stem cells signal transduction pathway].Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2008 Apr;22(4):492-5. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2008. PMID: 18575456 Review. Chinese.
Cited by
-
Notch signaling and cardiac repair.J Mol Cell Cardiol. 2012 Jun;52(6):1226-32. doi: 10.1016/j.yjmcc.2012.03.007. Epub 2012 Mar 21. J Mol Cell Cardiol. 2012. PMID: 22465038 Free PMC article. Review.
-
βA3/A1-crystallin and persistent fetal vasculature (PFV) disease of the eye.Biochim Biophys Acta. 2016 Jan;1860(1 Pt B):287-98. doi: 10.1016/j.bbagen.2015.05.017. Epub 2015 May 31. Biochim Biophys Acta. 2016. PMID: 26022148 Free PMC article. Review.
-
Astrocyte Differentiation of Human Pluripotent Stem Cells: New Tools for Neurological Disorder Research.Front Cell Neurosci. 2016 Sep 26;10:215. doi: 10.3389/fncel.2016.00215. eCollection 2016. Front Cell Neurosci. 2016. PMID: 27725795 Free PMC article. Review.
-
Isolation and characterization of GFAP-positive porcine neural stem/progenitor cells derived from a GFAP-CreERT2 transgenic piglet.BMC Vet Res. 2018 Nov 7;14(1):331. doi: 10.1186/s12917-018-1660-4. BMC Vet Res. 2018. PMID: 30404643 Free PMC article.
-
Notch signaling in glioblastoma: a developmental drug target?BMC Med. 2010 Nov 15;8:72. doi: 10.1186/1741-7015-8-72. BMC Med. 2010. PMID: 21078177 Free PMC article. Review.
References
-
- Androutsellis-Theotokis, A., R. R. Leker, F. Soldner, D. J. Hoeppner, R. Ravin, S. W. Poser, M. A. Rueger, S. K. Bae, R. Kittappa, and R. D. McKay. 2006. Notch signalling regulates stem cell numbers in vitro and in vivo. Nature 442:823-826. - PubMed
-
- Aster, J. C., E. S. Robertson, R. P. Hasserjian, J. R. Turner, E. Kieff, and J. Sklar. 1997. Oncogenic forms of NOTCH1 lacking either the primary binding site for RBP-Jκ or nuclear localization sequences retain the ability to associate with RBP-Jκ and activate transcription. J. Biol. Chem. 272:11336-11343. - PubMed
-
- Barnabe-Heider, F., J. A. Wasylnka, K. J. Fernandes, C. Porsche, M. Sendtner, D. R. Kaplan, and F. D. Miller. 2005. Evidence that embryonic neurons regulate the onset of cortical gliogenesis via cardiotrophin-1. Neuron 48:253-265. - PubMed
-
- Bertrand, N., D. S. Castro, and F. Guillemot. 2002. Proneural genes and the specification of neural cell types. Nat. Rev. Neurosci. 3:517-530. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous