

Identification of two protein kinases required for abscisic acid regulation of seed germination, root growth, and gene expression in Arabidopsis
- PMID: 17307925
- PMCID: PMC1867333
- DOI: 10.1105/tpc.106.048538
Identification of two protein kinases required for abscisic acid regulation of seed germination, root growth, and gene expression in Arabidopsis
Abstract
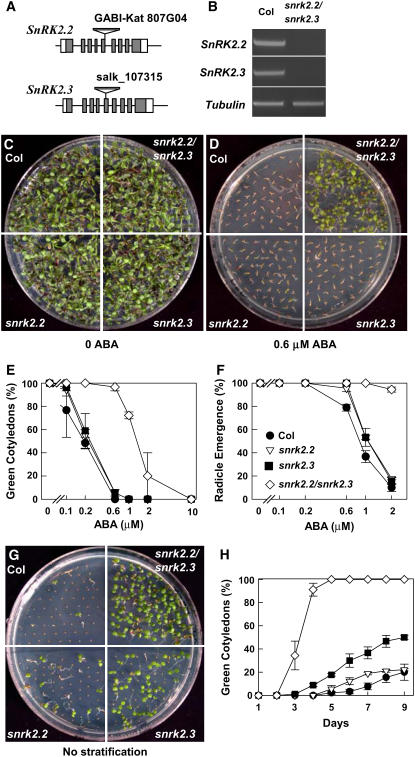
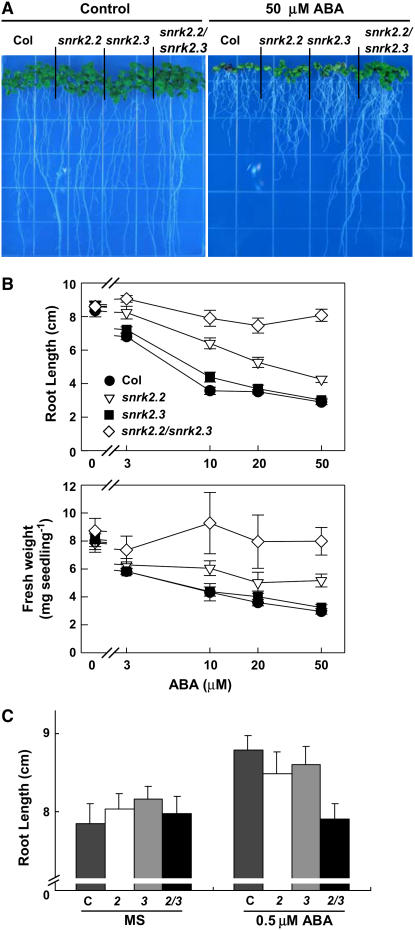
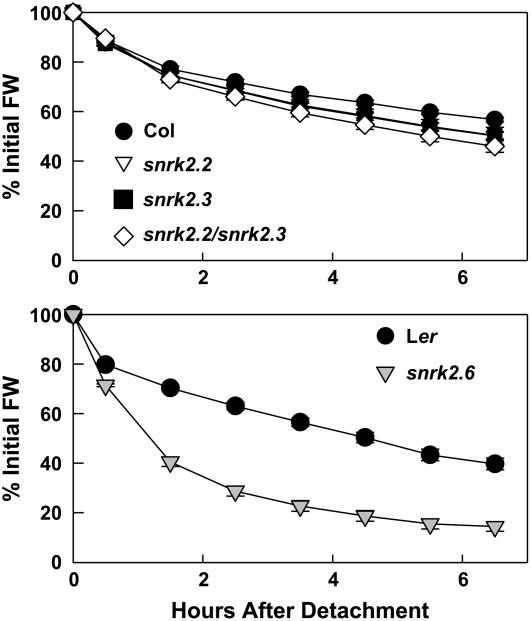
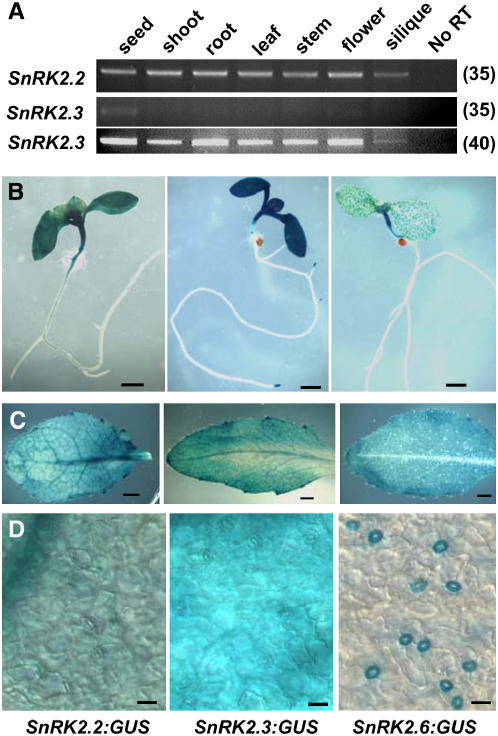
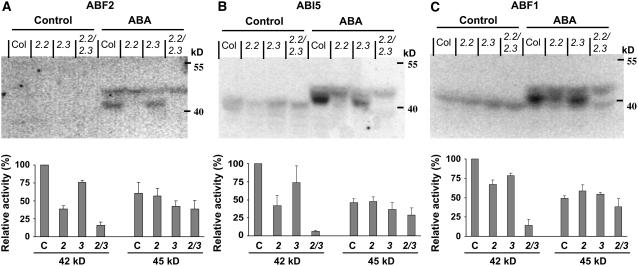
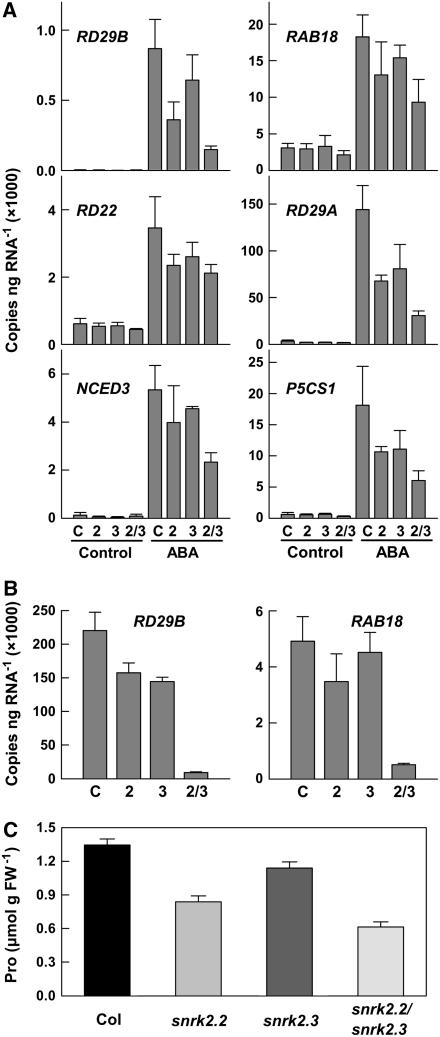
Abscisic acid (ABA) is an important phytohormone regulating various plant processes, including seed germination. Although phosphorylation has been suggested to be important, the protein kinases required for ABA signaling during seed germination and seedling growth remain elusive. Here, we show that two protein kinases, SNF1-RELATED PROTEIN KINASE2.2 (SnRK2.2) and SnRK2.3, control responses to ABA in seed germination, dormancy, and seedling growth in Arabidopsis thaliana. A snrk2.2 snrk2.3 double mutant, but not snrk2.2 or snrk2.3 single mutants, showed strong ABA-insensitive phenotypes in seed germination and root growth inhibition. Changes in seed dormancy and ABA-induced Pro accumulation consistent with ABA insensitivity were also observed. The snrk2.2 snrk2.3 double mutant had a greatly reduced level of a 42-kD kinase activity capable of phosphorylating peptides from ABF (for ABA Response Element Binding Factor) transcription factors. ABA-induced expression of several genes whose promoters contain an ABA response element (ABRE) was reduced in snrk2.2 snrk2.3, suggesting that the mechanism of SnRK2.2 and SnRK2.3 action in ABA signaling involves the activation of ABRE-driven gene expression through the phosphorylation of ABFs. Together, these results demonstrate that SnRK2.2 and SnRK2.3 are redundant but key protein kinases that mediate a major part of ABA signaling in Arabidopsis.
Figures
Similar articles
-
Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signaling are essential for the control of seed development and dormancy.Plant Cell Physiol. 2009 Jul;50(7):1345-63. doi: 10.1093/pcp/pcp083. Epub 2009 Jun 18. Plant Cell Physiol. 2009. PMID: 19541597
-
Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development.Plant J. 2014 Nov;80(4):654-68. doi: 10.1111/tpj.12670. Plant J. 2014. PMID: 25231920
-
WRKY41 controls Arabidopsis seed dormancy via direct regulation of ABI3 transcript levels not downstream of ABA.Plant J. 2014 Sep;79(5):810-23. doi: 10.1111/tpj.12597. Epub 2014 Jul 28. Plant J. 2014. PMID: 24946881
-
Pivotal role of the AREB/ABF-SnRK2 pathway in ABRE-mediated transcription in response to osmotic stress in plants.Physiol Plant. 2013 Jan;147(1):15-27. doi: 10.1111/j.1399-3054.2012.01635.x. Epub 2012 May 16. Physiol Plant. 2013. PMID: 22519646 Review.
-
Updates on the Role of ABSCISIC ACID INSENSITIVE 5 (ABI5) and ABSCISIC ACID-RESPONSIVE ELEMENT BINDING FACTORs (ABFs) in ABA Signaling in Different Developmental Stages in Plants.Cells. 2021 Aug 5;10(8):1996. doi: 10.3390/cells10081996. Cells. 2021. PMID: 34440762 Free PMC article. Review.
Cited by
-
Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress.Plant Cell Rep. 2013 Jul;32(7):985-1006. doi: 10.1007/s00299-013-1414-5. Epub 2013 Mar 19. Plant Cell Rep. 2013. PMID: 23508256 Review.
-
ABA signaling in stress-response and seed development.Plant Cell Rep. 2013 Jul;32(7):959-70. doi: 10.1007/s00299-013-1418-1. Epub 2013 Mar 28. Plant Cell Rep. 2013. PMID: 23535869 Review.
-
Arabidopsis PYR/PYL/RCAR receptors play a major role in quantitative regulation of stomatal aperture and transcriptional response to abscisic acid.Plant Cell. 2012 Jun;24(6):2483-96. doi: 10.1105/tpc.112.098574. Epub 2012 Jun 26. Plant Cell. 2012. PMID: 22739828 Free PMC article.
-
Molecular character of a phosphatase 2C (PP2C) gene relation to stress tolerance in Arabidopsis thaliana.Mol Biol Rep. 2013 Mar;40(3):2633-44. doi: 10.1007/s11033-012-2350-0. Epub 2012 Dec 26. Mol Biol Rep. 2013. PMID: 23268310 Free PMC article.
-
SUCROSE NONFERMENTING1-RELATED PROTEIN KINASE2.6, an ortholog of OPEN STOMATA1, is a negative regulator of strawberry fruit development and ripening.Plant Physiol. 2015 Mar;167(3):915-30. doi: 10.1104/pp.114.251314. Epub 2015 Jan 21. Plant Physiol. 2015. PMID: 25609556 Free PMC article.
References
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657. - PubMed
-
- Assmann, S.M. (2003). OPEN STOMATA1 opens the door to ABA signaling in Arabidopsis guard cells. Trends Plant Sci. 8 151–153. - PubMed
-
- Bates, L.S., Waldren, R.P., and Teare, I.D. (1973). Rapid determination of free proline in water-stress studies. Plant Soil 39 205–207.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
