

Involvement of maternal embryonic leucine zipper kinase (MELK) in mammary carcinogenesis through interaction with Bcl-G, a pro-apoptotic member of the Bcl-2 family
- PMID: 17280616
- PMCID: PMC1851384
- DOI: 10.1186/bcr1650
Involvement of maternal embryonic leucine zipper kinase (MELK) in mammary carcinogenesis through interaction with Bcl-G, a pro-apoptotic member of the Bcl-2 family
Abstract
Introduction: Cancer therapies directed at specific molecular targets in signaling pathways of cancer cells, such as tamoxifen, aromatase inhibitors and trastuzumab, have proven useful for treatment of advanced breast cancers. However, increased risk of endometrial cancer with long-term tamoxifen administration and of bone fracture due to osteoporosis in postmenopausal women undergoing aromatase inhibitor therapy are recognized side effects. These side effects as well as drug resistance make it necessary to search for novel molecular targets for drugs on the basis of well-characterized mechanisms of action.
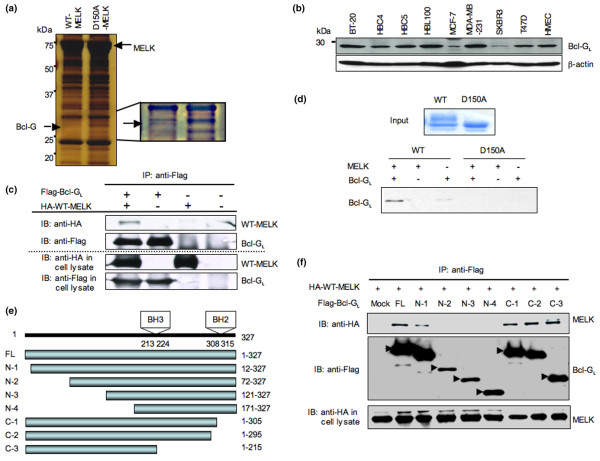
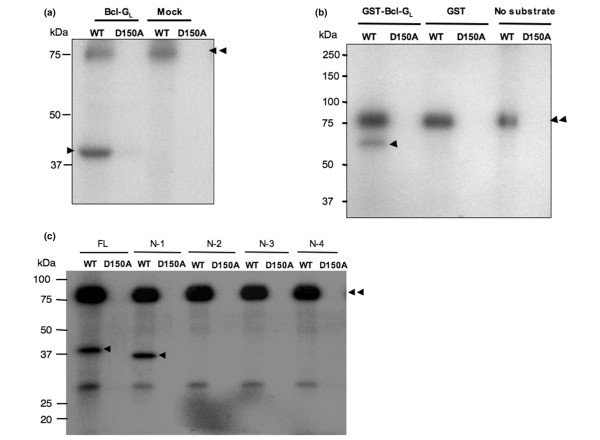
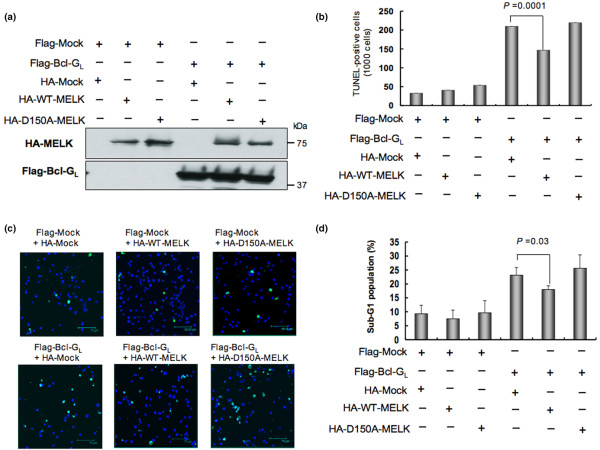
Methods: Using accurate genome-wide expression profiles of breast cancers, we found maternal embryonic leucine-zipper kinase (MELK) to be significantly overexpressed in the great majority of breast cancer cells. To assess whether MELK has a role in mammary carcinogenesis, we knocked down the expression of endogenous MELK in breast cancer cell lines using mammalian vector-based RNA interference. Furthermore, we identified a long isoform of Bcl-G (Bcl-GL), a pro-apoptotic member of the Bcl-2 family, as a possible substrate for MELK by pull-down assay with recombinant wild-type and kinase-dead MELK. Finally, we performed TUNEL assays and FACS analysis, measuring proportions of apoptotic cells, to investigate whether MELK is involved in the apoptosis cascade through the Bcl-GL-related pathway.
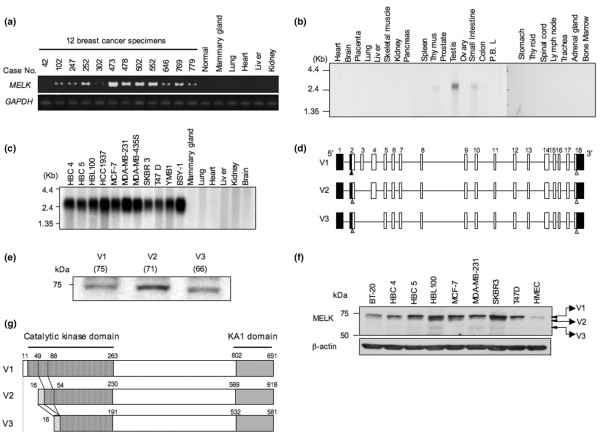
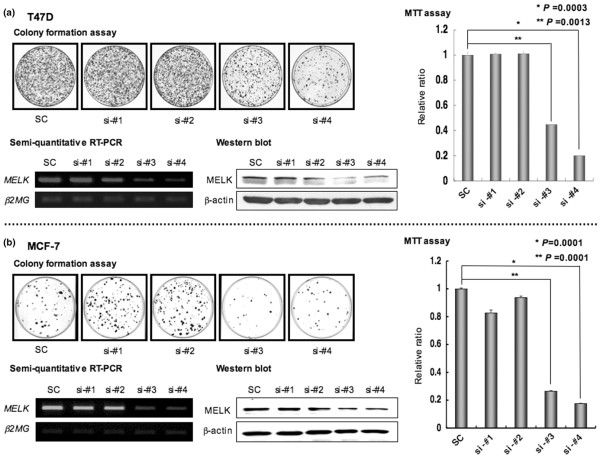
Results: Northern blot analyses on multiple human tissues and cancer cell lines demonstrated that MELK was overexpressed at a significantly high level in a great majority of breast cancers and cell lines, but was not expressed in normal vital organs (heart, liver, lung and kidney). Suppression of MELK expression by small interfering RNA significantly inhibited growth of human breast cancer cells. We also found that MELK physically interacted with Bcl-GL through its amino-terminal region. Immunocomplex kinase assay showed that Bcl-GL was specifically phosphorylated by MELK in vitro. TUNEL assays and FACS analysis revealed that overexpression of wild-type MELK suppressed Bcl-GL-induced apoptosis, while that of D150A-MELK did not.
Conclusion: Our findings suggest that the kinase activity of MELK is likely to affect mammary carcinogenesis through inhibition of the pro-apoptotic function of Bcl-GL. The kinase activity of MELK could be a promising molecular target for development of therapy for patients with breast cancers.
Figures
Similar articles
-
Maternal embryonic leucine zipper kinase/murine protein serine-threonine kinase 38 is a promising therapeutic target for multiple cancers.Cancer Res. 2005 Nov 1;65(21):9751-61. doi: 10.1158/0008-5472.CAN-04-4531. Cancer Res. 2005. PMID: 16266996
-
Putative tumor suppressor Lats2 induces apoptosis through downregulation of Bcl-2 and Bcl-x(L).Exp Cell Res. 2004 Aug 15;298(2):329-38. doi: 10.1016/j.yexcr.2004.04.031. Exp Cell Res. 2004. PMID: 15265683
-
Tumor-specific RNAi targeting eIF4E suppresses tumor growth, induces apoptosis and enhances cisplatin cytotoxicity in human breast carcinoma cells.Breast Cancer Res Treat. 2009 Feb;113(3):443-56. doi: 10.1007/s10549-008-9956-x. Epub 2008 Mar 10. Breast Cancer Res Treat. 2009. PMID: 18327707
-
Advancements of antisense oligonucleotides in treatment of breast cancer.Acta Pharmacol Sin. 2003 Apr;24(4):289-95. Acta Pharmacol Sin. 2003. PMID: 12676065 Review.
-
p53 and its downstream proteins as molecular targets of cancer.Mol Carcinog. 2006 Jun;45(6):409-15. doi: 10.1002/mc.20231. Mol Carcinog. 2006. PMID: 16652354 Review.
Cited by
-
A potential molecular model for studying apoptosis enhanced by the interaction of BCL-G with JAB1 in swine.Oncotarget. 2016 Sep 27;7(39):62912-62924. doi: 10.18632/oncotarget.11230. Oncotarget. 2016. PMID: 27542239 Free PMC article.
-
Maternal embryonic leucine zipper kinase serves as a poor prognosis marker and therapeutic target in osteosarcoma.Oncol Rep. 2020 Sep;44(3):1037-1048. doi: 10.3892/or.2020.7686. Epub 2020 Jul 13. Oncol Rep. 2020. PMID: 32705239 Free PMC article.
-
Targeting MELK in tumor cells and tumor microenvironment: from function and mechanism to therapeutic application.Clin Transl Oncol. 2025 Mar;27(3):887-900. doi: 10.1007/s12094-024-03664-5. Epub 2024 Aug 26. Clin Transl Oncol. 2025. PMID: 39187643 Review.
-
The Discovery of Novel Biomarkers Improves Breast Cancer Intrinsic Subtype Prediction and Reconciles the Labels in the METABRIC Data Set.PLoS One. 2015 Jul 1;10(7):e0129711. doi: 10.1371/journal.pone.0129711. eCollection 2015. PLoS One. 2015. PMID: 26132585 Free PMC article.
-
Thr55 phosphorylation of p21 by MPK38/MELK ameliorates defects in glucose, lipid, and energy metabolism in diet-induced obese mice.Cell Death Dis. 2019 May 16;10(6):380. doi: 10.1038/s41419-019-1616-z. Cell Death Dis. 2019. PMID: 31097688 Free PMC article.
References
-
- Navolanic PM, Mccubrey JA. Pharmacological breast cancer therapy (review) Int J Oncol. 2005;27:1341–1344. - PubMed
-
- Molina MA, Codony-Servat J, Albanell J, Rojo F, Arribas J, Baselga J. Trastuzumab (herceptin), a humanized anti-Her2 receptor monoclonal antibody, inhibits basal and activated Her2 ectodomain cleavage in breast cancer cells. Cancer Res. 2001;61:4744–4749. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
