

Genes within the Idd5 and Idd9/11 diabetes susceptibility loci affect the pathogenic activity of B cells in nonobese diabetic mice
- PMID: 17082619
- PMCID: PMC2886968
- DOI: 10.4049/jimmunol.177.10.7033
Genes within the Idd5 and Idd9/11 diabetes susceptibility loci affect the pathogenic activity of B cells in nonobese diabetic mice
Abstract
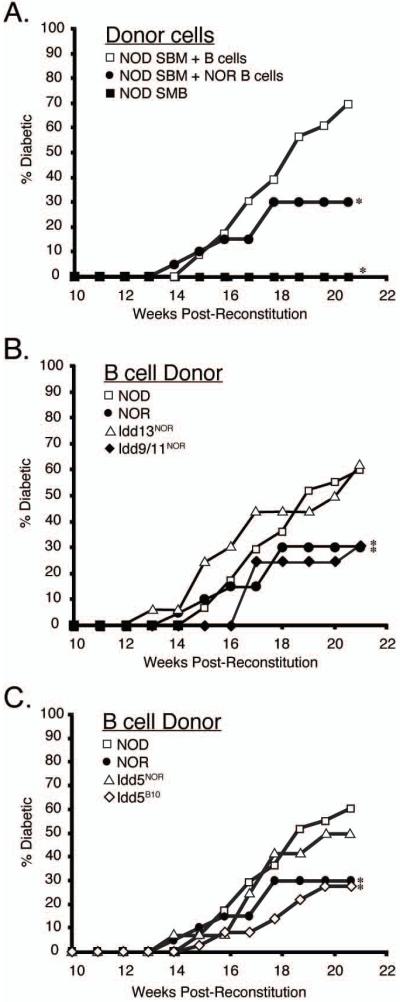
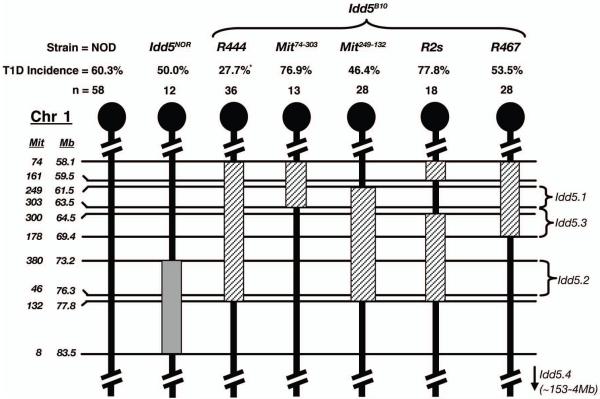
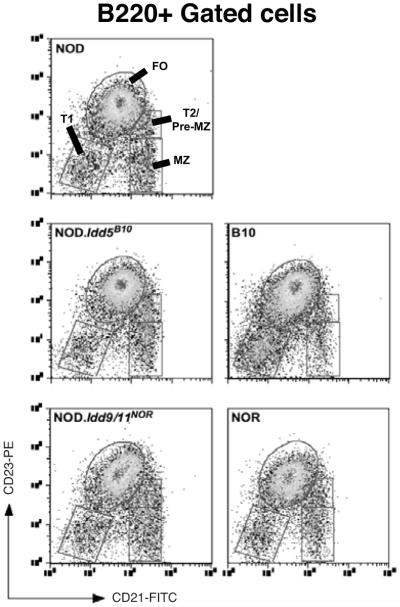
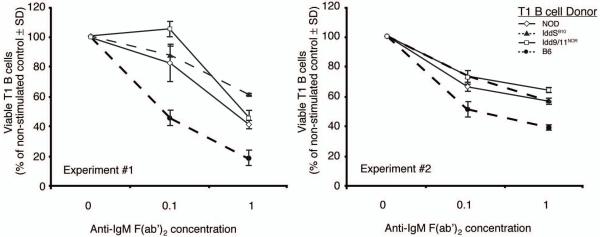
Autoreactive T cells clearly mediate the pancreatic beta cell destruction causing type 1 diabetes (T1D). However, studies in NOD mice indicate that B cells also contribute to pathogenesis because their ablation by introduction of an Igmunull mutation elicits T1D resistance. T1D susceptibility is restored in NOD.Igmunull mice that are irradiated and reconstituted with syngeneic bone marrow plus NOD B cells, but not syngeneic bone marrow alone. Thus, we hypothesized some non-MHC T1D susceptibility (Idd) genes contribute to disease by allowing development of pathogenic B cells. Supporting this hypothesis was the finding that unlike those from NOD donors, engraftment with B cells from H2g7 MHC-matched, but T1D-resistant, nonobese-resistant (NOR) mice failed to restore full disease susceptibility in NOD.Igmunull recipients. T1D resistance in NOR mice is mainly encoded within the Idd13, Idd5.2, and Idd9/11 loci. B cells from NOD congenic stocks containing Idd9/11 or Idd5.1/5.2-resistance loci, respectively, derived from the NOR or C57BL/10 strains were characterized by suppressed diabetogenic activity. Immature autoreactive B cells in NOD mice have an impaired ability to be rendered anergic upon Ag engagement. Interestingly, both Idd5.1/5.2 and Idd9/11-resistance loci were found to normalize this B cell tolerogenic process, which may represent a mechanism contributing to the inhibition of T1D.
Figures


Similar articles
-
Idd9/11 genetic locus regulates diabetogenic activity of CD4 T-cells in nonobese diabetic (NOD) mice.Diabetes. 2008 Dec;57(12):3273-80. doi: 10.2337/db08-0767. Epub 2008 Sep 5. Diabetes. 2008. PMID: 18776136 Free PMC article.
-
Subcongenic analyses reveal complex interactions between distal chromosome 4 genes controlling diabetogenic B cells and CD4 T cells in nonobese diabetic mice.J Immunol. 2012 Aug 1;189(3):1406-17. doi: 10.4049/jimmunol.1200120. Epub 2012 Jun 25. J Immunol. 2012. PMID: 22732593 Free PMC article.
-
Through regulation of TCR expression levels, an Idd7 region gene(s) interactively contributes to the impaired thymic deletion of autoreactive diabetogenic CD8+ T cells in nonobese diabetic mice.J Immunol. 2008 Mar 1;180(5):3250-9. doi: 10.4049/jimmunol.180.5.3250. J Immunol. 2008. PMID: 18292549
-
The importance of the Non Obese Diabetic (NOD) mouse model in autoimmune diabetes.J Autoimmun. 2016 Jan;66:76-88. doi: 10.1016/j.jaut.2015.08.019. Epub 2015 Sep 26. J Autoimmun. 2016. PMID: 26403950 Free PMC article. Review.
-
Gene-gene interactions in the NOD mouse model of type 1 diabetes.Adv Immunol. 2008;100:151-75. doi: 10.1016/S0065-2776(08)00806-7. Adv Immunol. 2008. PMID: 19111166 Review.
Cited by
-
Genome-wide transcriptional analyses of islet-specific CD4+ T cells identify Idd9 genes controlling diabetogenic T cell function.J Immunol. 2015 Mar 15;194(6):2654-63. doi: 10.4049/jimmunol.1401288. Epub 2015 Feb 11. J Immunol. 2015. PMID: 25672752 Free PMC article.
-
Idd9.1 locus controls the suppressive activity of FoxP3+CD4+CD25+ regulatory T-cells.Diabetes. 2010 Jan;59(1):272-81. doi: 10.2337/db09-0648. Epub 2009 Oct 15. Diabetes. 2010. PMID: 19833887 Free PMC article.
-
B Cell Receptor Affinity for Insulin Dictates Autoantigen Acquisition and B Cell Functionality in Autoimmune Diabetes.J Clin Med. 2016 Nov 8;5(11):98. doi: 10.3390/jcm5110098. J Clin Med. 2016. PMID: 27834793 Free PMC article.
-
Idd9/11 genetic locus regulates diabetogenic activity of CD4 T-cells in nonobese diabetic (NOD) mice.Diabetes. 2008 Dec;57(12):3273-80. doi: 10.2337/db08-0767. Epub 2008 Sep 5. Diabetes. 2008. PMID: 18776136 Free PMC article.
-
B cell depletion in autoimmune diabetes: insights from murine models.Expert Opin Ther Targets. 2011 Jun;15(6):703-14. doi: 10.1517/14728222.2011.561320. Epub 2011 Mar 3. Expert Opin Ther Targets. 2011. PMID: 21366498 Free PMC article. Review.
References
-
- Fox CJ, Danska JS. Independent genetic regulation of T-cell and antigen-presenting cell participation in autoimmune islet inflammation. Diabetes. 1998;47:331–338. - PubMed
-
- Jarpe AJ, Hickman MR, Anderson JT, Winter WE, Peck AB. Flow cytometric enumeration of mononuclear cell populations infiltrating the islets of Langer-hans in prediabetic NOD mice: development of a model of autoimmune insulitis for type I diabetes. Reg Immunol. 1990;3:305–317. - PubMed
-
- Gottlieb PA, Eisenbarth GS. Diagnosis and treatment of pre-insulin dependent diabetes. Annu Rev Med. 1998;49:391–405. - PubMed
-
- Noorchashm H, Noorchashm N, Kern J, Rostami SY, Barker CF, Naji A. B-cells are required for the initiation of insulitis and sialitis in nonobese diabetic mice. Diabetes. 1997;46:941–946. - PubMed
-
- Serreze DV, Chapman HD, Varnum DS, Hanson MS, Reifsnyder PC, Richard SD, Fleming SA, Leiter EH, Shultz LD. B lymphocytes are essential for the initiation of T-cell-mediated autoimmune diabetes: analysis of a new “speed congenic” stock of NOD.Ig mu null mice. J Exp Med. 1996;184:2049–2053. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials