

EC5S ubiquitin complex is recruited by KSHV latent antigen LANA for degradation of the VHL and p53 tumor suppressors
- PMID: 17069461
- PMCID: PMC1626105
- DOI: 10.1371/journal.ppat.0020116
EC5S ubiquitin complex is recruited by KSHV latent antigen LANA for degradation of the VHL and p53 tumor suppressors
Abstract
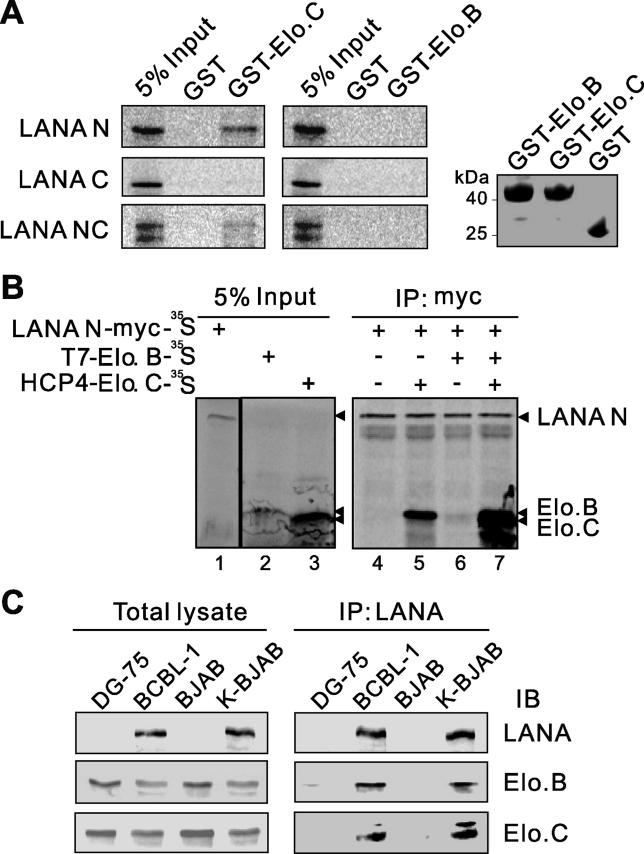
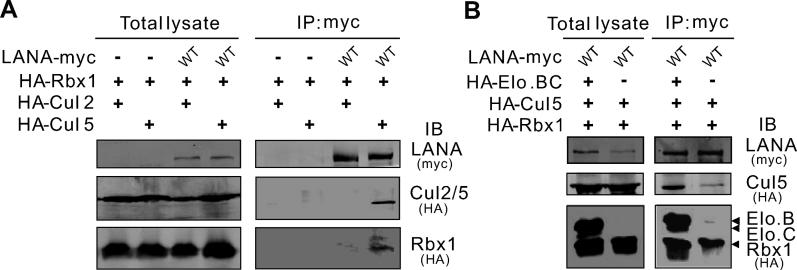
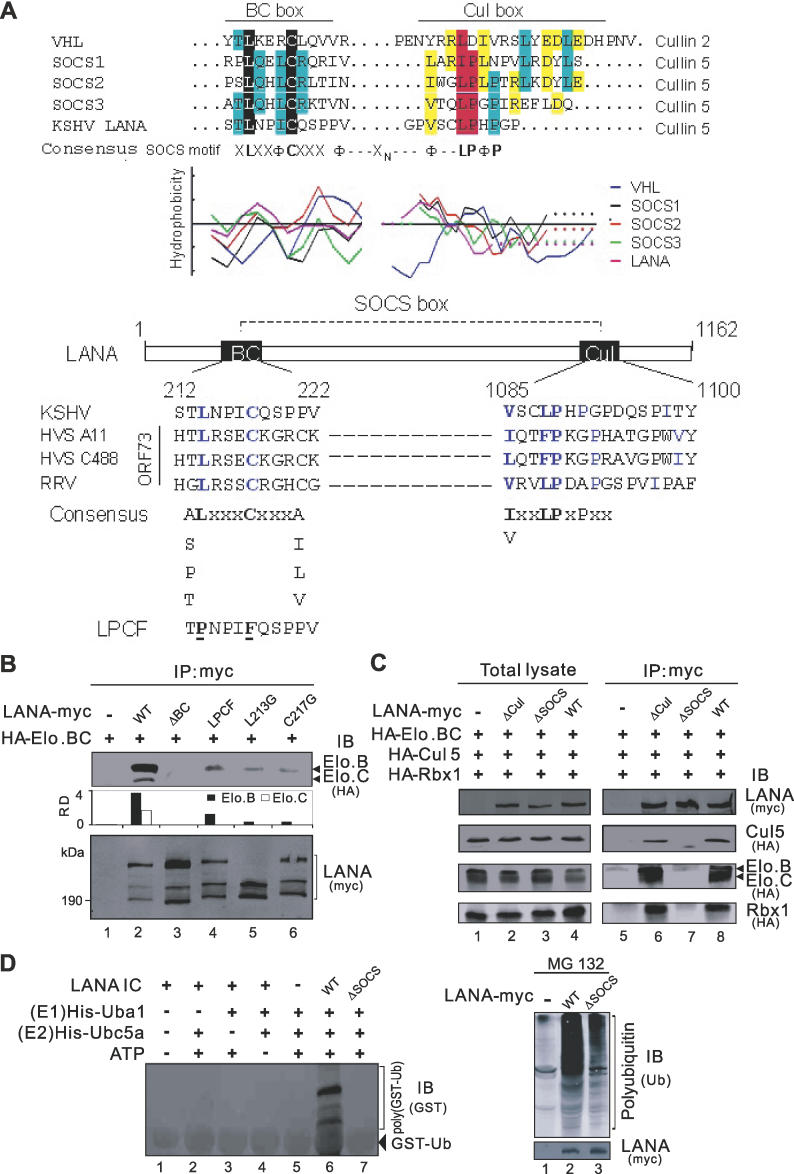
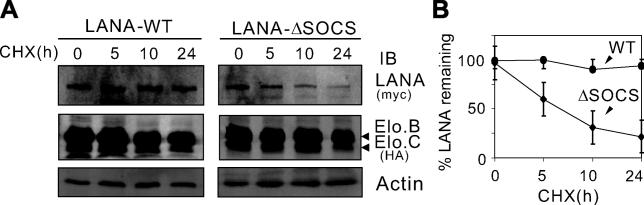
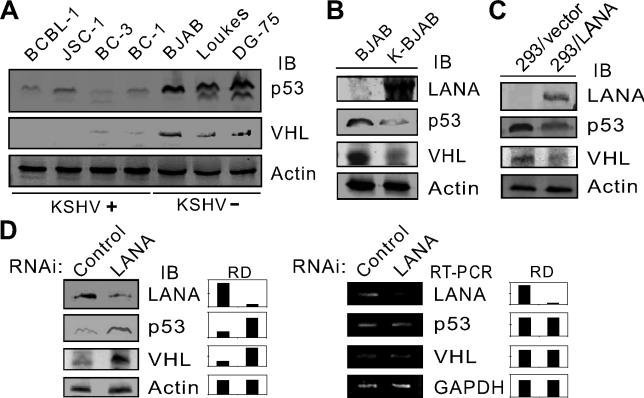
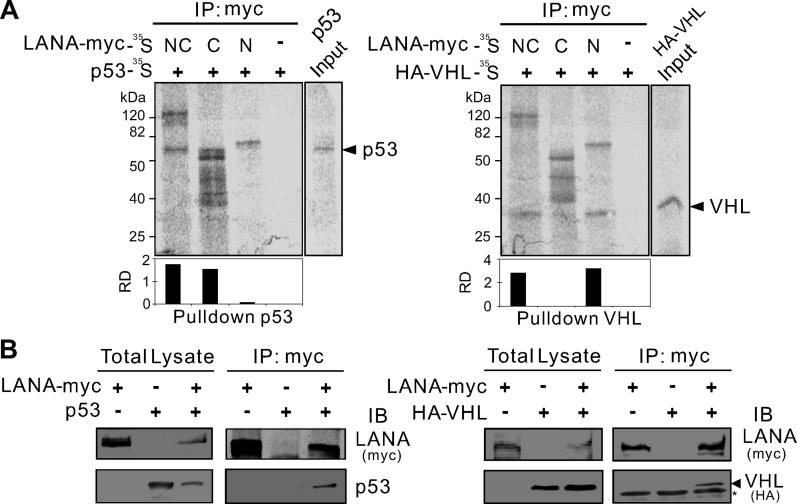
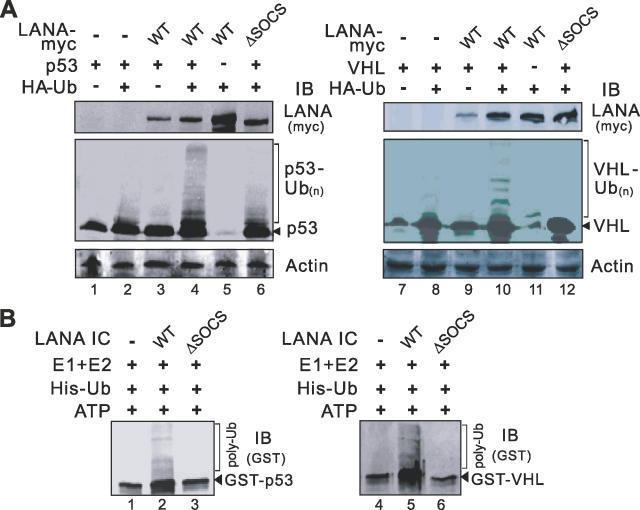
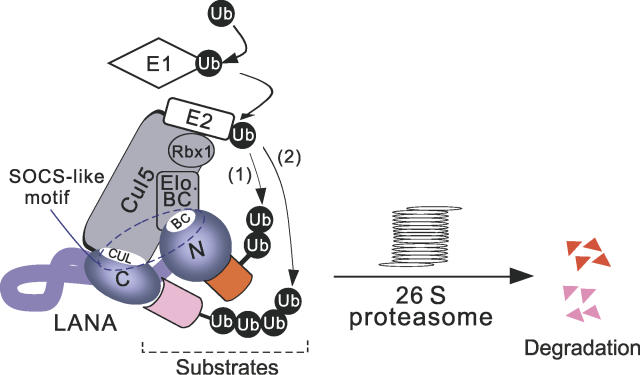
Cellular protein degradation pathways can be utilized by viruses to establish an environment that favors their propagation. Here we report that the Kaposi's sarcoma-associated herpesvirus (KSHV)-encoded latency-associated nuclear antigen (LANA) directly functions as a component of the EC5S ubiquitin complex targeting the tumor suppressors von Hippel-Lindau (VHL) and p53 for degradation. We have characterized a suppressor of cytokine signaling box-like motif within LANA composed of an Elongin B and C box and a Cullin box, which is spatially located at its amino and carboxyl termini. This motif is necessary for LANA interaction with the Cul5-Elongin BC complex, to promote polyubiquitylation of cellular substrates VHL and p53 in vitro via its amino- and carboxyl-terminal binding domain, respectively. In transfected cells as well as KSHV-infected B lymphoma cells, LANA expression stimulates degradation of VHL and p53. Additionally, specific RNA interference-mediated LANA knockdown stabilized VHL and p53 in primary effusion lymphoma cells. Thus, manipulation of tumor suppressors by LANA potentially provides a favorable environment for progression of KSHV-infected tumor cells.
Conflict of interest statement
Figures
Similar articles
-
A potential alpha-helix motif in the amino terminus of LANA encoded by Kaposi's sarcoma-associated herpesvirus is critical for nuclear accumulation of HIF-1alpha in normoxia.J Virol. 2007 Oct;81(19):10413-23. doi: 10.1128/JVI.00611-07. Epub 2007 Jul 18. J Virol. 2007. PMID: 17634230 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus-encoded LANA positively affects on ubiquitylation of p53.Biochem Biophys Res Commun. 2010 Dec 10;403(2):194-7. doi: 10.1016/j.bbrc.2010.11.004. Epub 2010 Nov 5. Biochem Biophys Res Commun. 2010. PMID: 21056547
-
Kaposi's Sarcoma-Associated Herpesvirus LANA Modulates the Stability of the E3 Ubiquitin Ligase RLIM.J Virol. 2020 Feb 14;94(5):e01578-19. doi: 10.1128/JVI.01578-19. Print 2020 Feb 14. J Virol. 2020. PMID: 31801865 Free PMC article.
-
Role of SOCS and VHL Proteins in Neuronal Differentiation and Development.Int J Mol Sci. 2023 Feb 15;24(4):3880. doi: 10.3390/ijms24043880. Int J Mol Sci. 2023. PMID: 36835292 Free PMC article. Review.
-
Do VHL and HIF-1 mirror p53 and Mdm-2? Degradation-transactivation loops of oncoproteins and tumor suppressors.Oncogene. 2001 Jan 18;20(3):395-8. doi: 10.1038/sj.onc.1204055. Oncogene. 2001. PMID: 11313969 Review.
Cited by
-
VHL negatively regulates SARS coronavirus replication by modulating nsp16 ubiquitination and stability.Biochem Biophys Res Commun. 2015 Apr 3;459(2):270-276. doi: 10.1016/j.bbrc.2015.02.097. Epub 2015 Feb 28. Biochem Biophys Res Commun. 2015. PMID: 25732088 Free PMC article.
-
Hypoxic reactivation of Kaposi's sarcoma associated herpesvirus.Cell Insight. 2024 Sep 7;3(6):100200. doi: 10.1016/j.cellin.2024.100200. eCollection 2024 Dec. Cell Insight. 2024. PMID: 39391006 Free PMC article. Review.
-
KSHV reactivation and novel implications of protein isomerization on lytic switch control.Viruses. 2015 Jan 12;7(1):72-109. doi: 10.3390/v7010072. Viruses. 2015. PMID: 25588053 Free PMC article. Review.
-
Targeting Kaposi's sarcoma associated herpesvirus encoded protease (ORF17) by a lysophosphatidic acid molecule for treating KSHV associated diseases.Front Cell Dev Biol. 2023 Jan 17;11:1060156. doi: 10.3389/fcell.2023.1060156. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 36733461 Free PMC article.
-
SOCS and Herpesviruses, With Emphasis on Cytomegalovirus Retinitis.Front Immunol. 2019 Apr 11;10:732. doi: 10.3389/fimmu.2019.00732. eCollection 2019. Front Immunol. 2019. PMID: 31031749 Free PMC article. Review.
References
-
- Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J, et al. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi's sarcoma. Science. 1994;266:1865–1869. - PubMed
-
- Boshoff C, Schulz TF, Kennedy MM, Graham AK, Fisher C, et al. Kaposi's sarcoma-associated herpesvirus infects endothelial and spindle cells. Nat Med. 1995;1:1274–1278. - PubMed
-
- Arvanitakis L, Mesri EA, Nador RG, Said JW, Asch AS, et al. Establishment and characterization of a primary effusion (body cavity-based) lymphoma cell line (BC-3) harboring Kaposi's sarcoma-associated herpesvirus (KSHV/HHV-8) in the absence of Epstein-Barr virus. Blood. 1996;88:2648–2654. - PubMed
-
- Hayward GS. Human herpesvirus 8 latent-state gene expression and apoptosis in Kaposi's sarcoma lesions. J Natl Cancer Inst. 1999;91:1705–1707. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
