

pH-Dependence of extrinsic and intrinsic H(+)-ion mobility in the rat ventricular myocyte, investigated using flash photolysis of a caged-H(+) compound
- PMID: 17056723
- PMCID: PMC1751406
- DOI: 10.1529/biophysj.106.096560
pH-Dependence of extrinsic and intrinsic H(+)-ion mobility in the rat ventricular myocyte, investigated using flash photolysis of a caged-H(+) compound
Abstract
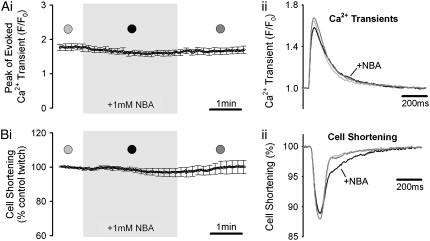
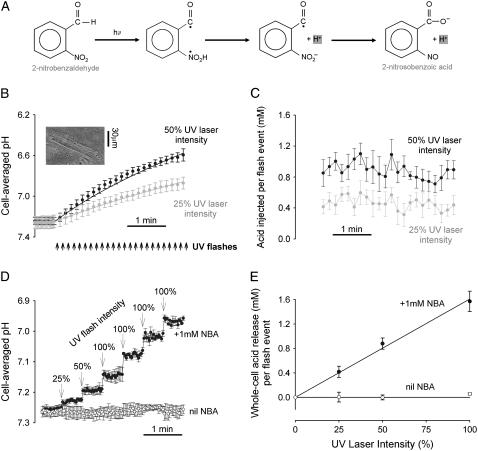
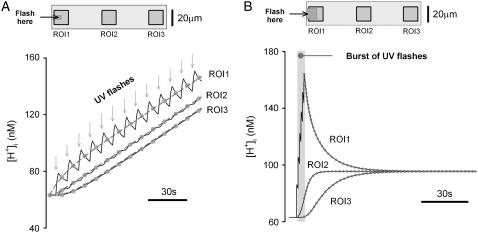
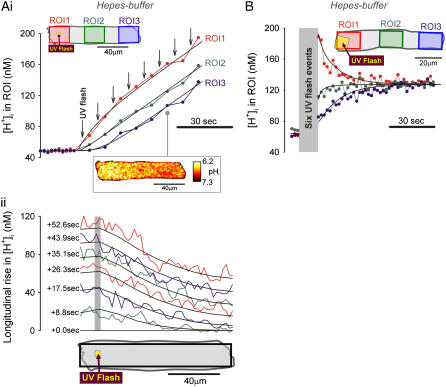
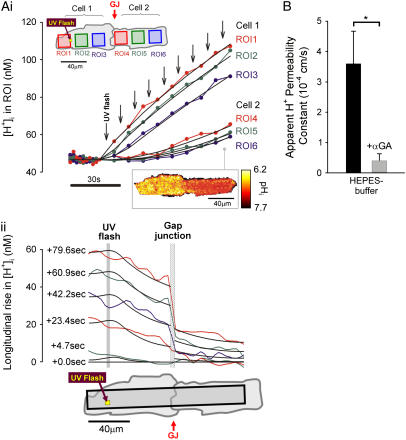
Passive H(+)-ion mobility within eukaryotic cells is low, due to H(+)-ion binding to cytoplasmic buffers. A localized intracellular acidosis can therefore persist for seconds or even minutes. Because H(+)-ions modulate so many biological processes, spatial intracellular pH (pH(i))-regulation becomes important for coordinating cellular activity. We have investigated spatial pH(i)-regulation in single and paired ventricular myocytes from rat heart by inducing a localized intracellular acid-load, while confocally imaging pH(i) using the pH-fluorophore, carboxy-SNARF-1. We present a novel method for localizing the acid-load. This involves the intracellular photolytic uncaging of H(+)-ions from a membrane-permeant acid-donor, 2-nitrobenzaldehyde. The subsequent spatial pH(i)-changes are consistent with intracellular H(+)-mobility and cell-to-cell H(+)-permeability constants measured using more conventional acid-loading techniques. We use the method to investigate the effect of reducing pH(i) on intrinsic (non-CO(2)/HCO(3)(-) buffer-dependent) and extrinsic (CO(2)/HCO(3)(-) buffer-dependent) components of H(i)(+)-mobility. We find that although both components mediate spatial regulation of pH within the cell, their ability to do so declines sharply at low pH(i). Thus acidosis severely slows intracellular H(+)-ion movement. This can result in spatial pH(i) nonuniformity, particularly during the stimulation of sarcolemmal Na(+)-H(+) exchange. Intracellular acidosis thus presents a window of vulnerability in the spatial coordination of cellular function.
Figures

Similar articles
-
Spatial regulation of intracellular pH in multicellular strands of neonatal rat cardiomyocytes.Cardiovasc Res. 2010 Mar 1;85(4):729-38. doi: 10.1093/cvr/cvp343. Epub 2009 Oct 14. Cardiovasc Res. 2010. PMID: 19828673
-
Spatial regulation of intracellular pH in the ventricular myocyte.Ann N Y Acad Sci. 2005 Jun;1047:271-82. doi: 10.1196/annals.1341.024. Ann N Y Acad Sci. 2005. PMID: 16093503
-
Relationship between intracellular pH and proton mobility in rat and guinea-pig ventricular myocytes.J Physiol. 2005 Aug 1;566(Pt 3):793-806. doi: 10.1113/jphysiol.2005.086165. Epub 2005 May 26. J Physiol. 2005. PMID: 15919716 Free PMC article.
-
Spatial aspects of intracellular pH regulation in heart muscle.Prog Biophys Mol Biol. 2006 Jan-Apr;90(1-3):207-24. doi: 10.1016/j.pbiomolbio.2005.06.004. Epub 2005 Jul 7. Prog Biophys Mol Biol. 2006. PMID: 16125756 Review.
-
Modelling intracellular H(+) ion diffusion.Prog Biophys Mol Biol. 2003 Oct;83(2):69-100. doi: 10.1016/s0079-6107(03)00027-0. Prog Biophys Mol Biol. 2003. PMID: 12865074 Review.
Cited by
-
Cytosolic pH buffering during exercise and recovery in skeletal muscle of patients with McArdle's disease.Eur J Appl Physiol. 2009 Mar;105(5):687-94. doi: 10.1007/s00421-008-0950-0. Epub 2008 Dec 9. Eur J Appl Physiol. 2009. PMID: 19066935
-
Regional acidosis locally inhibits but remotely stimulates Ca2+ waves in ventricular myocytes.Cardiovasc Res. 2017 Jul 1;113(8):984-995. doi: 10.1093/cvr/cvx033. Cardiovasc Res. 2017. PMID: 28339694 Free PMC article.
-
Voltage-gated proton channels: what's next?J Physiol. 2008 Nov 15;586(22):5305-24. doi: 10.1113/jphysiol.2008.161703. Epub 2008 Sep 18. J Physiol. 2008. PMID: 18801839 Free PMC article. Review.
-
Coupled Ca2+/H+ transport by cytoplasmic buffers regulates local Ca2+ and H+ ion signaling.Proc Natl Acad Sci U S A. 2013 May 28;110(22):E2064-73. doi: 10.1073/pnas.1222433110. Epub 2013 May 15. Proc Natl Acad Sci U S A. 2013. PMID: 23676270 Free PMC article.
-
Facilitation by intracellular carbonic anhydrase of Na+ -HCO3- co-transport but not Na+ / H+ exchange activity in the mammalian ventricular myocyte.J Physiol. 2014 Mar 1;592(5):991-1007. doi: 10.1113/jphysiol.2013.265439. Epub 2013 Dec 2. J Physiol. 2014. PMID: 24297849 Free PMC article.
References
-
- Ellis, D., and R. C. Thomas. 1976. Microelectrode measurement of the intracellular pH of mammalian heart cells. Nature. 262:224–225. - PubMed
-
- Komukai, K., F. Brette, C. Pascarel, and C. H. Orchard. 2002. Electrophysiological response of rat ventricular myocytes to acidosis. Am. J. Physiol. Heart Circ. Physiol. 283:H412–H422. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
