

COPI activity coupled with fatty acid biosynthesis is required for viral replication
- PMID: 17040126
- PMCID: PMC1599761
- DOI: 10.1371/journal.ppat.0020102
COPI activity coupled with fatty acid biosynthesis is required for viral replication
Abstract
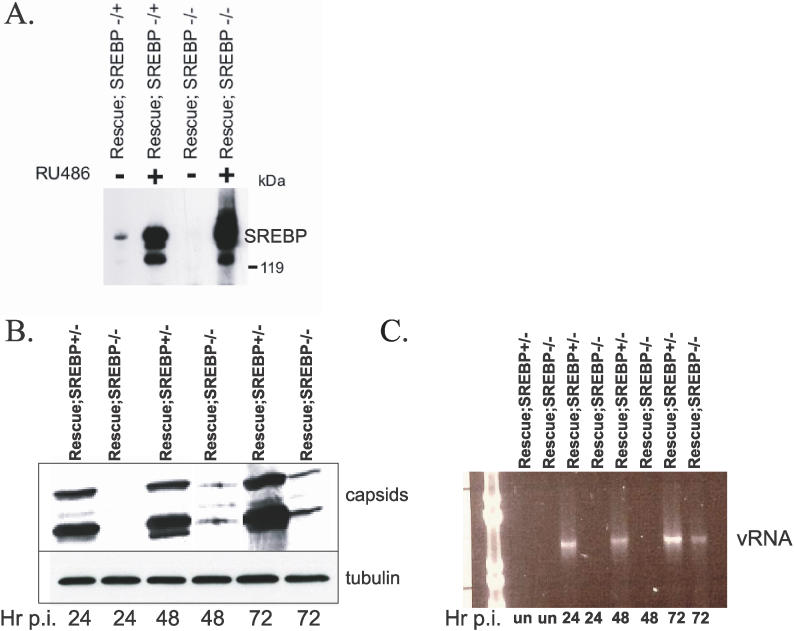
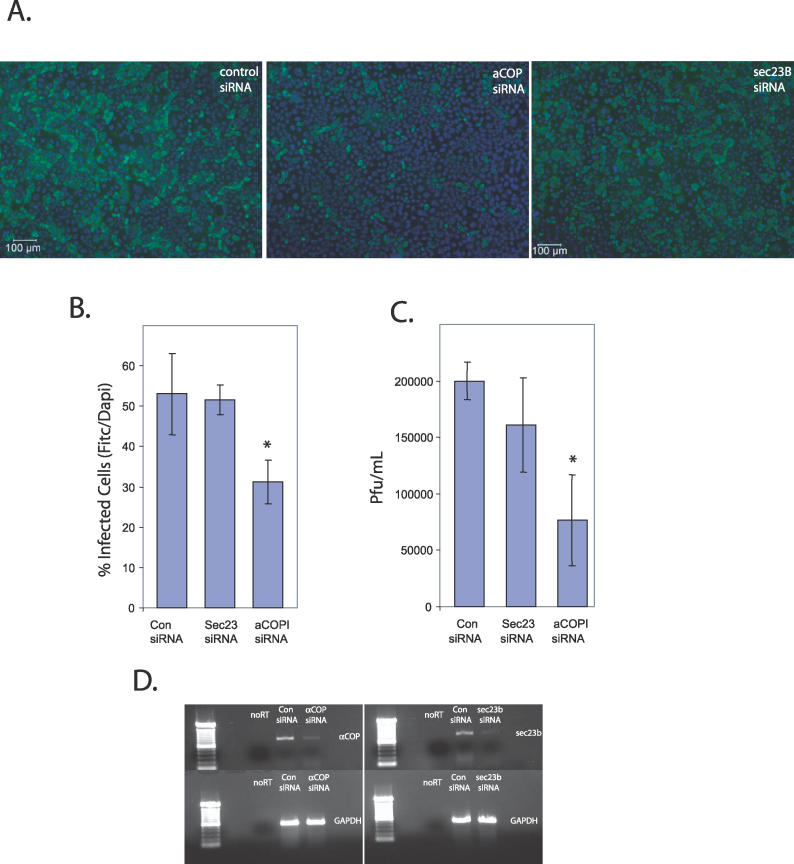
During infection by diverse viral families, RNA replication occurs on the surface of virally induced cytoplasmic membranes of cellular origin. How this process is regulated, and which cellular factors are required, has been unclear. Moreover, the host-pathogen interactions that facilitate the formation of this new compartment might represent critical determinants of viral pathogenesis, and their elucidation may lead to novel insights into the coordination of vesicular trafficking events during infection. Here we show that in Drosophila cells, Drosophila C virus remodels the Golgi apparatus and forms a novel vesicular compartment, on the surface of which viral RNA replication takes place. Using genome-wide RNA interference screening, we found that this step in the viral lifecycle requires at least two host encoded pathways: the coat protein complex I (COPI) coatamer and fatty acid biosynthesis. Our results integrate, clarify, and extend numerous observations concerning the cell biology of viral replication, allowing us to conclude that the coupling of new cellular membrane formation with the budding of these vesicles from the Golgi apparatus allows for the regulated generation of this new virogenic organelle, which is essential for viral replication. Additionally, because these pathways are also limiting in flies and in human cells infected with the related RNA virus poliovirus, they may represent novel targets for antiviral therapies.
Conflict of interest statement
Figures
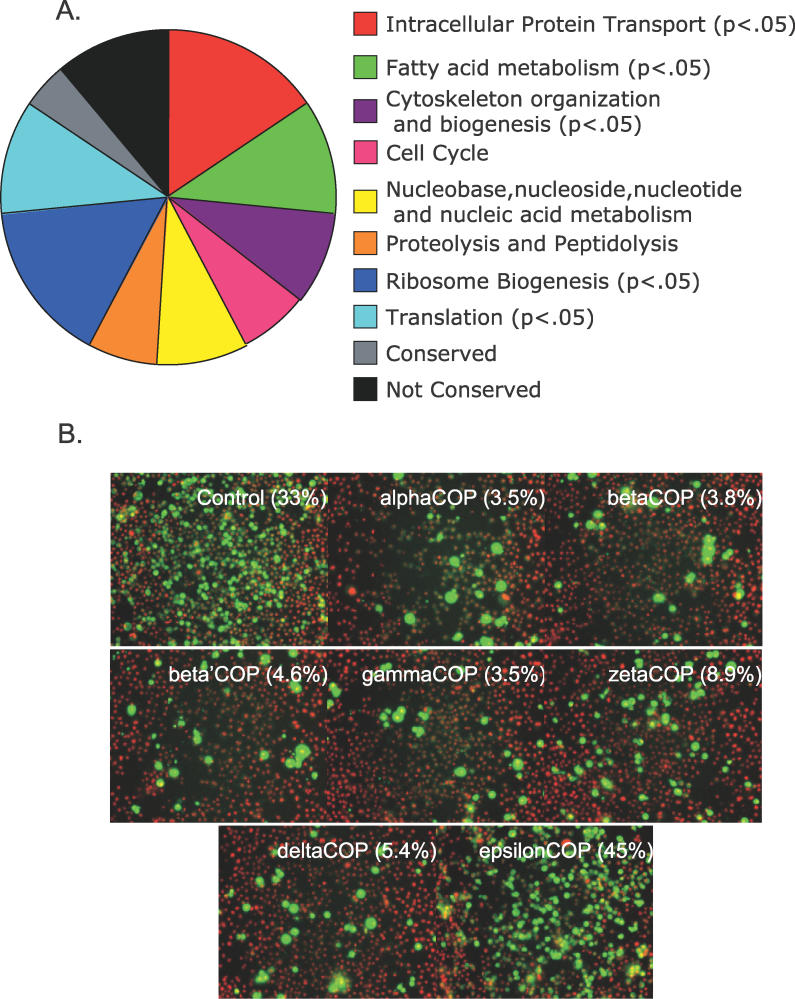
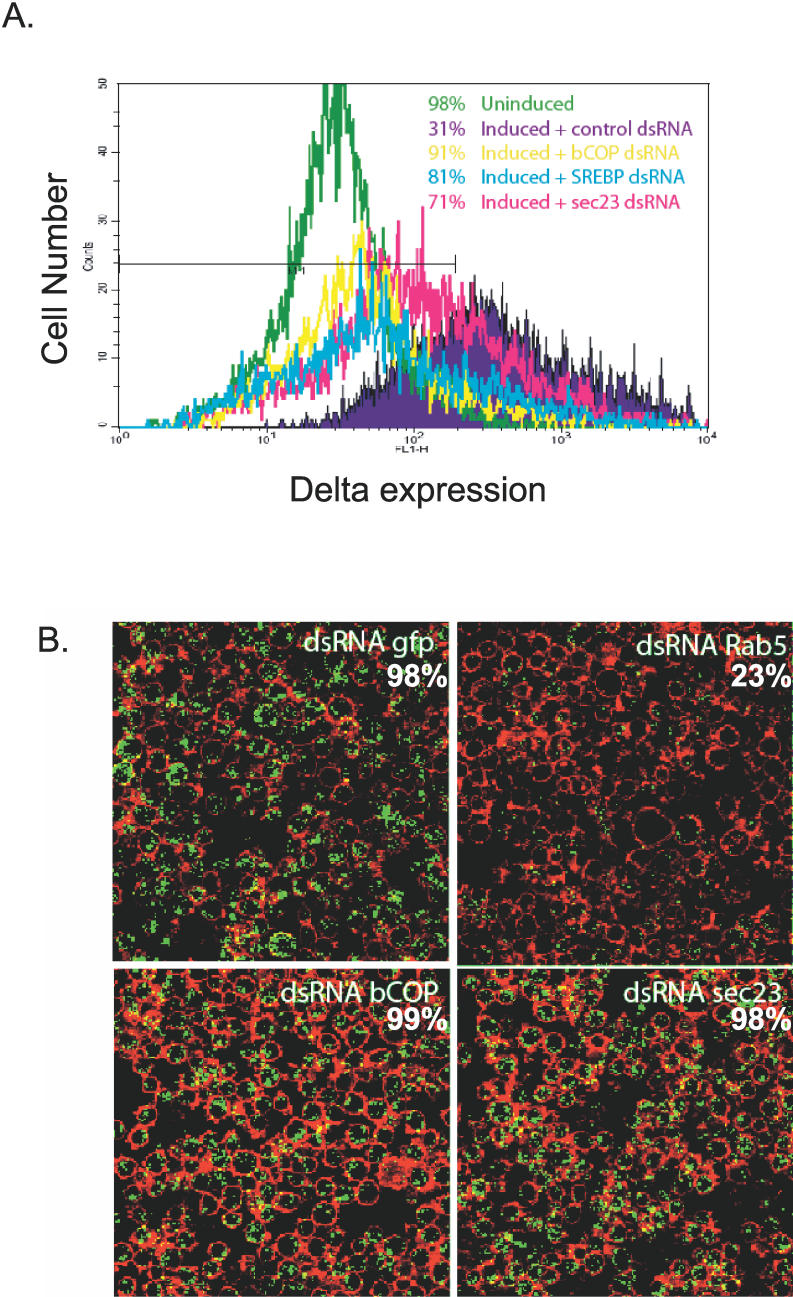
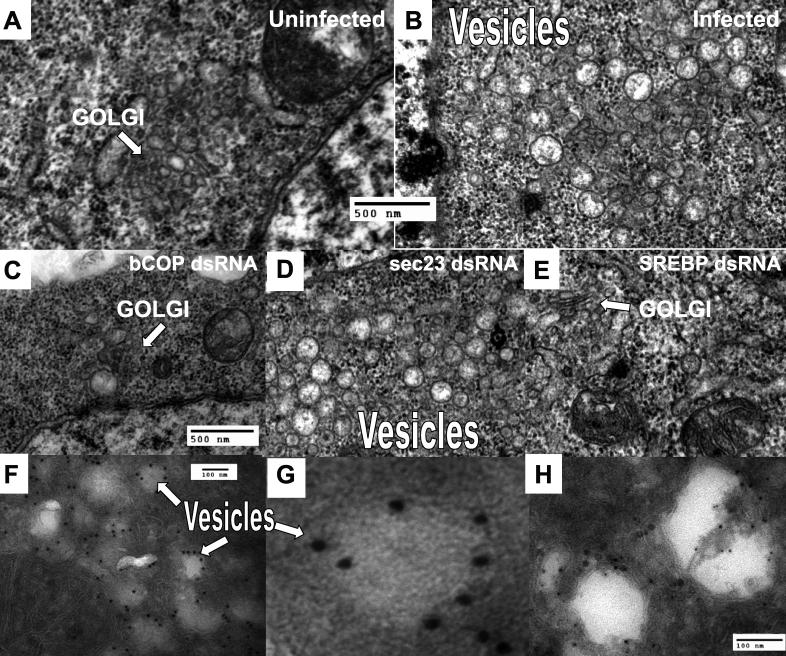
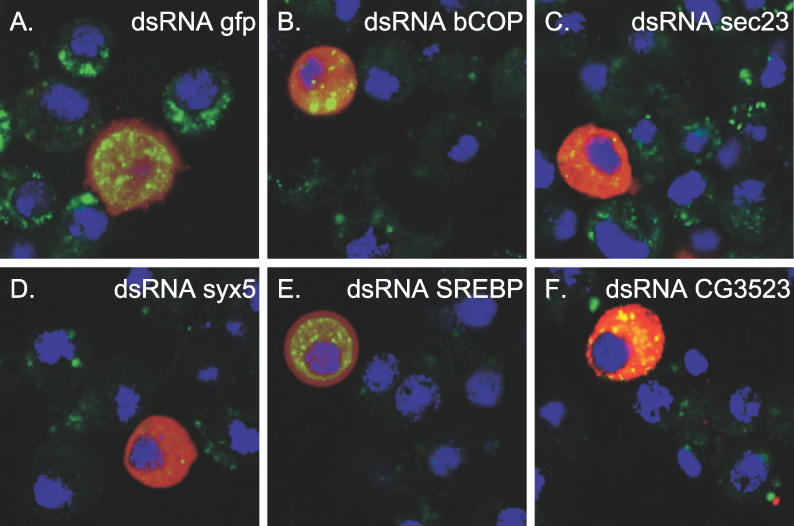
Similar articles
-
COPI is required for enterovirus 71 replication.PLoS One. 2012;7(5):e38035. doi: 10.1371/journal.pone.0038035. Epub 2012 May 25. PLoS One. 2012. PMID: 22662263 Free PMC article.
-
Generation of unique poliovirus RNA replication organelles.mBio. 2014 Feb 25;5(2):e00833-13. doi: 10.1128/mBio.00833-13. mBio. 2014. PMID: 24570367 Free PMC article.
-
Molecular characterization of Drosophila cells persistently infected with Flock House virus.Virology. 2011 Oct 10;419(1):43-53. doi: 10.1016/j.virol.2011.08.002. Epub 2011 Aug 26. Virology. 2011. PMID: 21872290 Free PMC article.
-
[Roles of COPI related proteins during virus replication].Bing Du Xue Bao. 2012 Mar;28(2):185-9. Bing Du Xue Bao. 2012. PMID: 22519182 Review. Chinese.
-
The Role of Electron Microscopy in Studying the Continuum of Changes in Membranous Structures during Poliovirus Infection.Viruses. 2015 Oct 12;7(10):5305-18. doi: 10.3390/v7102874. Viruses. 2015. PMID: 26473912 Free PMC article. Review.
Cited by
-
Lipids at the interface of virus-host interactions.Curr Opin Microbiol. 2012 Aug;15(4):512-8. doi: 10.1016/j.mib.2012.05.013. Epub 2012 Jun 9. Curr Opin Microbiol. 2012. PMID: 22682978 Free PMC article. Review.
-
Manipulation or capitulation: virus interactions with autophagy.Microbes Infect. 2012 Feb;14(2):126-39. doi: 10.1016/j.micinf.2011.09.007. Epub 2011 Oct 24. Microbes Infect. 2012. PMID: 22051604 Free PMC article. Review.
-
RNAi screening in mammalian cells to identify novel host cell molecules involved in the regulation of viral infections.Methods Mol Biol. 2011;721:397-405. doi: 10.1007/978-1-61779-037-9_25. Methods Mol Biol. 2011. PMID: 21431700 Free PMC article.
-
A key role for heat shock protein 70 in the localization and insertion of tombusvirus replication proteins to intracellular membranes.J Virol. 2009 Apr;83(7):3276-87. doi: 10.1128/JVI.02313-08. Epub 2009 Jan 19. J Virol. 2009. PMID: 19153242 Free PMC article.
-
Tombusviruses upregulate phospholipid biosynthesis via interaction between p33 replication protein and yeast lipid sensor proteins during virus replication in yeast.Virology. 2014 Dec;471-473:72-80. doi: 10.1016/j.virol.2014.10.005. Epub 2014 Oct 28. Virology. 2014. PMID: 25461533 Free PMC article.
References
-
- Lyle JM, Bullitt E, Bienz K, Kirkegaard K. Visualization and functional analysis of RNA-dependent RNA polymerase lattices. Science. 2002;296:2218–2222. - PubMed
-
- Jackson WT, Giddings TH, Jr, Taylor MP, Mulinyawe S, Rabinovitch M, et al. Subversion of cellular autophagosomal machinery by RNA viruses. PLoS Biol. 2005. e156. DOI: 10.1371/journal.pbio.0030156. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
