

Replication fork blockage by transcription factor-DNA complexes in Escherichia coli
- PMID: 17000639
- PMCID: PMC1636447
- DOI: 10.1093/nar/gkl682
Replication fork blockage by transcription factor-DNA complexes in Escherichia coli
Abstract
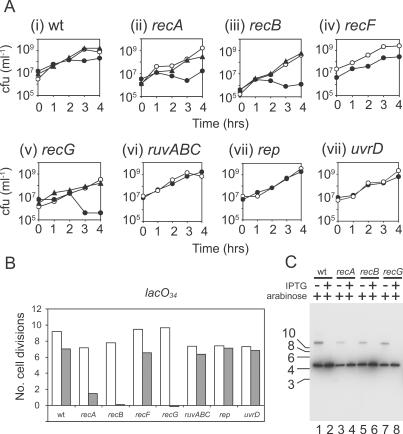
All organisms require mechanisms that resuscitate replication forks when they break down, reflecting the complex intracellular environments within which DNA replication occurs. Here we show that as few as three lac repressor-operator complexes block Escherichia coli replication forks in vitro regardless of the topological state of the DNA. Blockage with tandem repressor-operator complexes was also observed in vivo, demonstrating that replisomes have a limited ability to translocate through high affinity protein-DNA complexes. However, cells could tolerate tandem repressor-bound operators within the chromosome that were sufficient to block all forks in vitro. This discrepancy between in vitro and in vivo observations was at least partly explained by the ability of RecA, RecBCD and RecG to abrogate the effects of repressor-operator complexes on cell viability. However, neither RuvABC nor RecF were needed for normal cell growth in the face of such complexes. Holliday junction resolution by RuvABC and facilitated loading of RecA by RecF were not therefore critical for tolerance of protein-DNA blocks. We conclude that there is a trade-off between efficient genome duplication and other aspects of DNA metabolism such as transcriptional control, and that recombination enzymes, either directly or indirectly, provide the means to tolerate such conflicts.
Figures
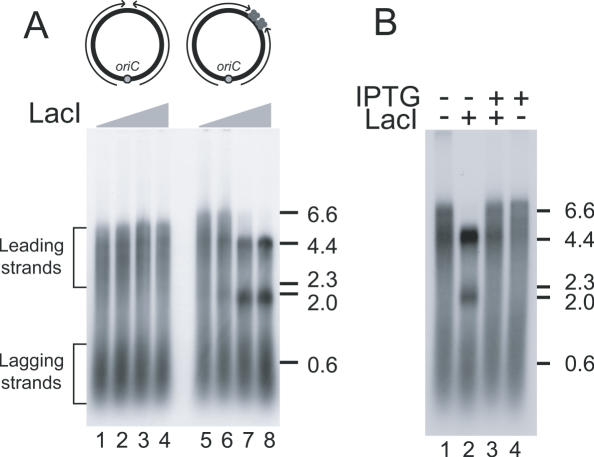
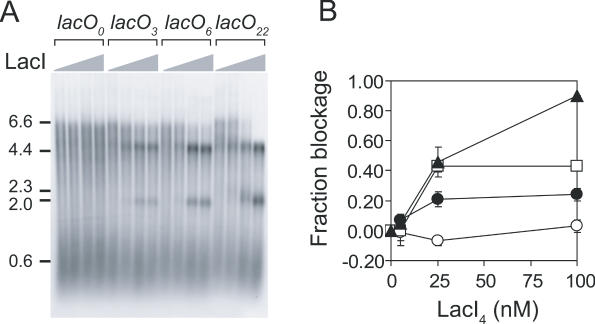
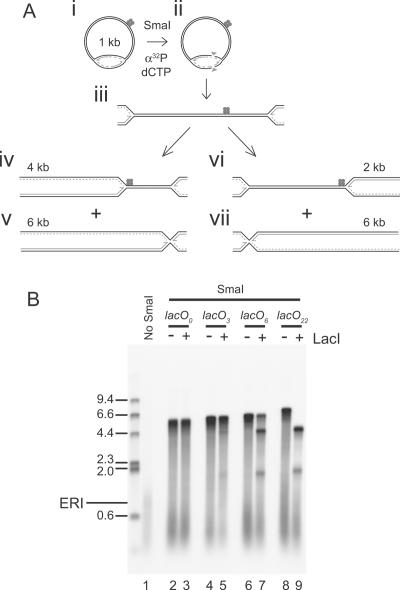
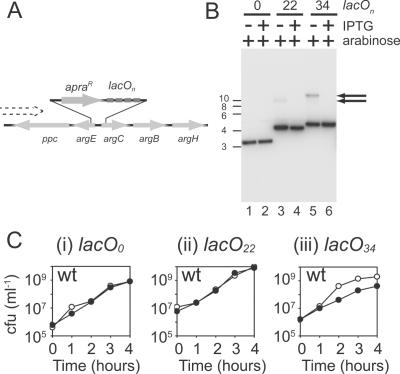

Similar articles
-
Replication forks blocked by protein-DNA complexes have limited stability in vitro.J Mol Biol. 2008 Aug 29;381(2):249-55. doi: 10.1016/j.jmb.2008.05.053. Epub 2008 May 29. J Mol Biol. 2008. PMID: 18602646
-
RuvABC-dependent double-strand breaks in dnaBts mutants require recA.Mol Microbiol. 2000 Nov;38(3):565-74. doi: 10.1046/j.1365-2958.2000.02152.x. Mol Microbiol. 2000. PMID: 11069680
-
Tracking of controlled Escherichia coli replication fork stalling and restart at repressor-bound DNA in vivo.EMBO J. 2006 Jun 7;25(11):2596-604. doi: 10.1038/sj.emboj.7601155. Epub 2006 May 25. EMBO J. 2006. PMID: 16724111 Free PMC article.
-
Single-molecule insight into stalled replication fork rescue in Escherichia coli.Nucleic Acids Res. 2021 May 7;49(8):4220-4238. doi: 10.1093/nar/gkab142. Nucleic Acids Res. 2021. PMID: 33744948 Free PMC article. Review.
-
What happens when replication and transcription complexes collide?Cell Cycle. 2010 Jul 1;9(13):2537-43. doi: 10.4161/cc.9.13.12122. Cell Cycle. 2010. PMID: 20581460 Free PMC article. Review.
Cited by
-
Recombination and replication.Cold Spring Harb Perspect Biol. 2014 Oct 23;6(11):a016550. doi: 10.1101/cshperspect.a016550. Cold Spring Harb Perspect Biol. 2014. PMID: 25341919 Free PMC article. Review.
-
Control of DNA replication in vitro using a reversible replication barrier.Nat Protoc. 2024 Jul;19(7):1940-1983. doi: 10.1038/s41596-024-00977-1. Epub 2024 Apr 9. Nat Protoc. 2024. PMID: 38594502 Free PMC article. Review.
-
The Functional Consequences of Eukaryotic Topoisomerase 1 Interaction with G-Quadruplex DNA.Genes (Basel). 2020 Feb 12;11(2):193. doi: 10.3390/genes11020193. Genes (Basel). 2020. PMID: 32059547 Free PMC article. Review.
-
RAD18, WRNIP1 and ATMIN promote ATM signalling in response to replication stress.Oncogene. 2016 Jul 28;35(30):4009-19. doi: 10.1038/onc.2015.427. Epub 2015 Nov 9. Oncogene. 2016. PMID: 26549024 Free PMC article.
-
Stimulation of UvrD helicase by UvrAB.J Biol Chem. 2009 Apr 3;284(14):9612-23. doi: 10.1074/jbc.M808030200. Epub 2009 Feb 10. J Biol Chem. 2009. PMID: 19208629 Free PMC article.
References
-
- Lindahl T. The Croonian Lecture, 1996: endogenous damage to DNA. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1996;351:1529–1538. - PubMed
-
- Rothstein R., Michel B., Gangloff S. Replication fork pausing and recombination or ‘gimme a break’. Genes Dev. 2000;14:1–10. - PubMed
-
- Mulcair M.D., Schaeffer P.M., Oakley A.J., Cross H.F., Neylon C., Hill T.M., Dixon N.E. A molecular mousetrap determines polarity of termination of DNA replication in E.coli. Cell. 2006;125:1309–1319. - PubMed
-
- Cox M.M., Goodman M.F., Kreuzer K.N., Sherratt D.J., Sandler S.J., Marians K.J. The importance of repairing stalled replication forks. Nature. 2000;404:37–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
