

Toll-like receptors differentially regulate CC and CXC chemokines in skeletal muscle via NF-kappaB and calcineurin
- PMID: 16982839
- PMCID: PMC1698076
- DOI: 10.1128/IAI.00286-06
Toll-like receptors differentially regulate CC and CXC chemokines in skeletal muscle via NF-kappaB and calcineurin
Abstract
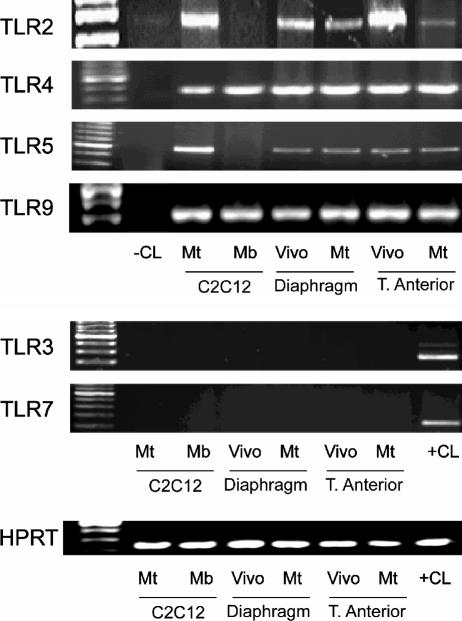
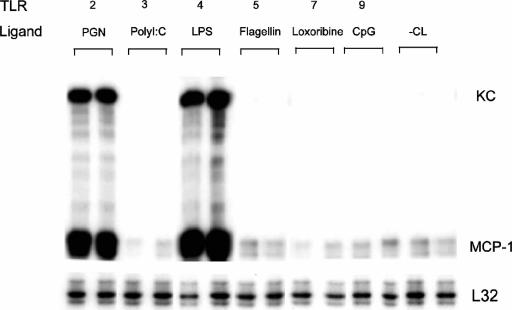
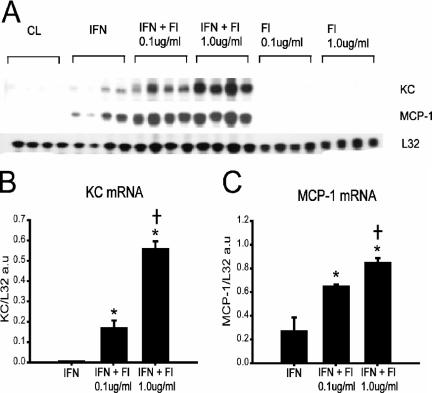
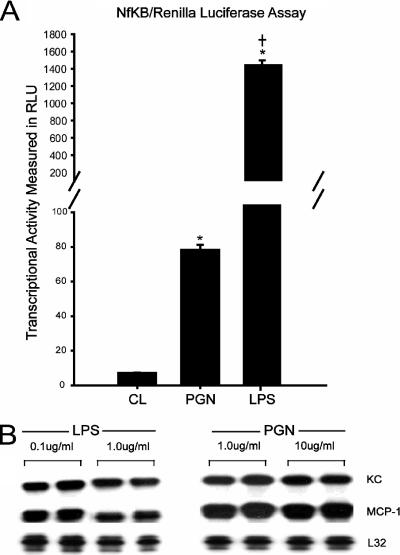
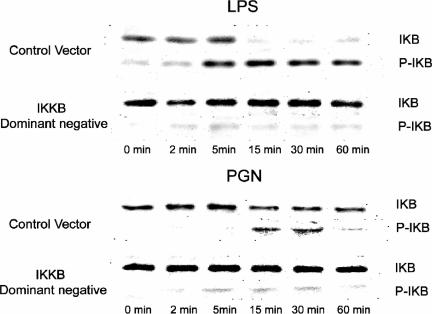
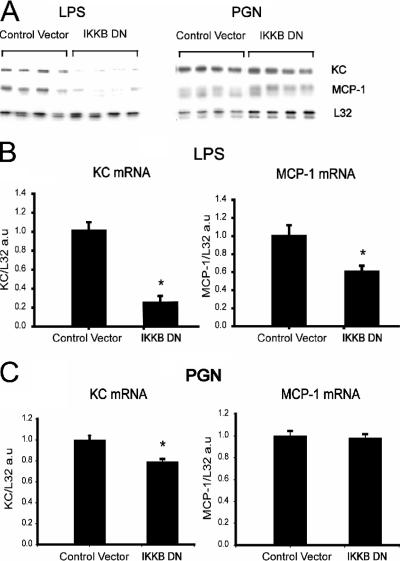
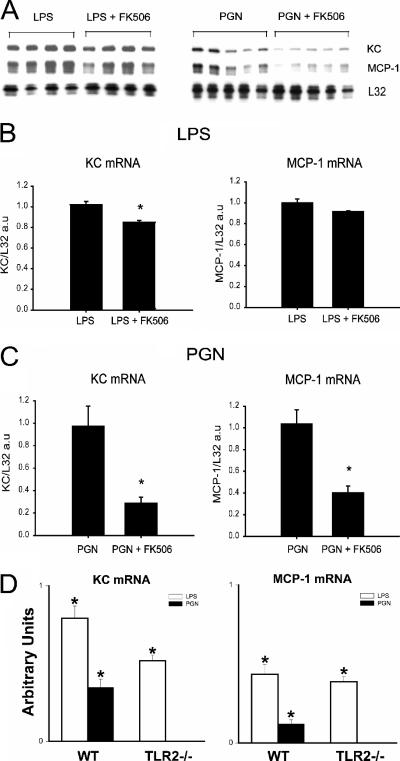
Immunologically active molecules such as cytokines and chemokines have been implicated in skeletal muscle weakness during sepsis as well as recovery from muscle injury. In sepsis, Toll-like receptors (TLRs) act as key sentinel molecules of the innate immune system. Here we determined skeletal muscle cell responses of two prototypical CC and CXC chemokine genes (monocyte chemoattractant protein 1 [MCP-1] and KC, respectively), to stimulation with specific TLR ligands. In addition, we examined whether NF-kappaB and calcineurin signaling are involved in these responses. Differentiated myotubes and intact whole muscles expressed TLR2, TLR4, TLR5, and TLR9. Stimulation with ligands for TLR2 (peptidoglycan) or TLR4 (LPS) elicited robust and equivalent levels of MCP-1 and KC mRNA expression, whereas stimulation of TLR5 (by flagellin) required gamma interferon priming to induce similar effects. Although both TLR2 and TLR4 ligands activated the NF-kappaB pathway, NF-kappaB reporter activity was approximately 20-fold greater after TLR4 stimulation than after TLR2 stimulation. Inhibitory effects of NF-kappaB blockade on TLR-mediated chemokine gene expression, by either pharmacological (pyrrolidine dithiocarbamate) or molecular (IKKbeta dominant-negative transfection) methods, were also more pronounced during TLR4 stimulation. In contrast, inhibitory effects on TLR-mediated chemokine expression of calcineurin blockade (by FK506) were greater for TLR2 than for TLR4 stimulation. MCP-1 and KC mRNA levels also demonstrated differential responses to NF-kappaB and calcineurin blockade during stimulation with specific TLR ligands. We conclude that skeletal muscle cells differentially utilize the NF-kappaB and calcineurin pathways in a TLR-specific manner to enable complex regulation of CC and CXC chemokine gene expression.
Figures

Similar articles
-
Endotoxin-induced chemokine expression in murine peritoneal mesothelial cells: the role of toll-like receptor 4.J Am Soc Nephrol. 2004 May;15(5):1289-99. J Am Soc Nephrol. 2004. PMID: 15100369
-
Ligand-induced differential cross-regulation of Toll-like receptors 2, 4 and 5 in intestinal epithelial cells.Mol Immunol. 2007 Jul;44(15):3702-14. doi: 10.1016/j.molimm.2007.04.001. Epub 2007 May 9. Mol Immunol. 2007. PMID: 17493681
-
Toll-like receptor 2, 3, and 4 expression and function in human airway smooth muscle.J Allergy Clin Immunol. 2006 Sep;118(3):641-8. doi: 10.1016/j.jaci.2006.05.013. Epub 2006 Jul 3. J Allergy Clin Immunol. 2006. PMID: 16950283
-
Signaling to NF-kappaB by Toll-like receptors.Trends Mol Med. 2007 Nov;13(11):460-9. doi: 10.1016/j.molmed.2007.09.002. Epub 2007 Oct 29. Trends Mol Med. 2007. PMID: 18029230 Review.
-
The role of Toll-like receptors in the regulation of neutrophil migration, activation, and apoptosis.Clin Infect Dis. 2005 Nov 15;41 Suppl 7:S421-6. doi: 10.1086/431992. Clin Infect Dis. 2005. PMID: 16237641 Review.
Cited by
-
MyD88 and Type I interferon receptor-mediated chemokine induction and monocyte recruitment during Listeria monocytogenes infection.J Immunol. 2009 Jul 15;183(2):1271-8. doi: 10.4049/jimmunol.0900460. Epub 2009 Jun 24. J Immunol. 2009. PMID: 19553532 Free PMC article.
-
Progressive Skeletal Muscle Atrophy in Muscular Dystrophies: A Role for Toll-like Receptor-Signaling in Disease Pathogenesis.Int J Mol Sci. 2020 Jun 22;21(12):4440. doi: 10.3390/ijms21124440. Int J Mol Sci. 2020. PMID: 32580419 Free PMC article. Review.
-
TLR5, a novel and unidentified inflammatory mediator in rheumatoid arthritis that correlates with disease activity score and joint TNF-α levels.J Immunol. 2012 Jul 1;189(1):475-83. doi: 10.4049/jimmunol.1102977. Epub 2012 Jun 1. J Immunol. 2012. PMID: 22661088 Free PMC article.
-
Pro-inflammatory mediation of myoblast proliferation.PLoS One. 2014 Mar 19;9(3):e92363. doi: 10.1371/journal.pone.0092363. eCollection 2014. PLoS One. 2014. PMID: 24647690 Free PMC article.
-
Enrichment of Newly Synthesized Proteins following treatment of C2C12 Myotubes with Endotoxin and Interferon-γ.Inflammation. 2022 Jun;45(3):1313-1331. doi: 10.1007/s10753-022-01622-3. Epub 2022 Jan 14. Inflammation. 2022. PMID: 35028803 Free PMC article.
References
-
- Abraham, S., T. Sweet, B. E. Sawaya, J. Rappaport, K. Khalili, and S. Amini. 2005. Cooperative interaction of C/EBP beta and Tat modulates MCP-1 gene transcription in astrocytes. J. Neuroimmunol. 160:219-227. - PubMed
-
- Alzuherri, H., and K. C. Chang. 2003. Calcineurin activates NF-kappaB in skeletal muscle C2C12 cells. Cell. Signal. 15:471-478. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
