

doi: 10.1128/JVI.00640-06.
Regulation of the transcription and replication cycle of human cytomegalovirus is insensitive to genetic elimination of the cognate NF-kappaB binding sites in the enhancer
Affiliations
- PMID: 16973595
- PMCID: PMC1617225
- DOI: 10.1128/JVI.00640-06
Item in Clipboard
Regulation of the transcription and replication cycle of human cytomegalovirus is insensitive to genetic elimination of the cognate NF-kappaB binding sites in the enhancer
J Virol.
2006 Oct.
Abstract
The role of NF-kappaB in regulating human cytomegalovirus (HCMV) replication and gene transcription remains controversial. Multiple, functional NF-kappaB response elements exist in the major immediate-early promoter (MIEP) enhancer of HCMV, suggesting a possible requirement for this transcription factor in lytic viral replication. Here we demonstrate by generating and analyzing HCMVs with alterations in the MIEP-enhancer that, although this region is essential for HCMV growth, none of the four NF-kappaB response elements contained within the enhancer are required for MIE gene expression or HCMV replication in multiple cell types. These data reveal the robustness of the regulatory network controlling the MIEP enhancer.
Figures
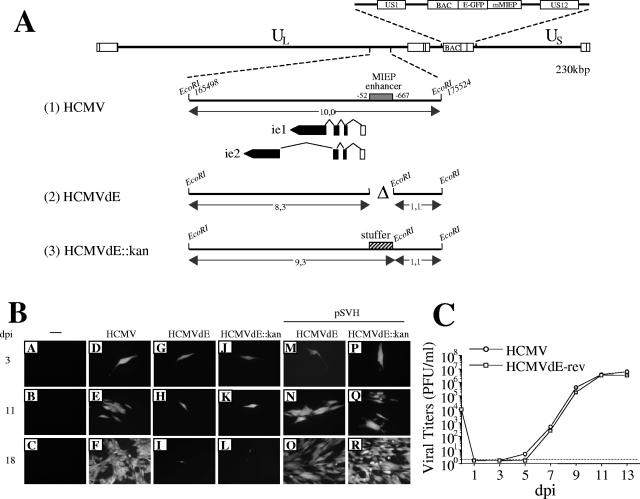
Effect of a deletion of the entire MIEP enhancer on HCMV. (A) Schematic representation of the enhancerless HCMV BAC genomes constructed. The top line represents the map of the parental HCMV BAC genome with the US1-US12 region expanded above, indicating the BAC sequences and the E-GFP gene under control of the MCMV MIEP (mMIEP) that replace the US2-US11 region. Below, the EcoRI J fragment (nucleotide sequences from 165498 to 175524 of the HCMV genome [28]) encompassing the HCMV MIE region is enlarged (line 1), with the structures of the ie1 and ie2 transcripts indicated. The first noncoding exon of the ie1/ie2 transcription unit is depicted as an open rectangle, and coding exons are shown solid. The shaded box marks the HCMV MIEP enhancer (extending from nucleotides −52 to −667 relative to the ie1/ie2 HCMV transcription start site). The two enhancerless HCMV BAC genomes, HCMVdE and HCMVdE::Kan, were derived from the parental HCMV BAC by the ET BAC mutagenesis method (5, 6, 24). The deletion of the entire HCMV enhancer (nucleotide sequences from 174713 to 175328 of the HCMV genome) in HCMVdE (line 2) is indicated by a “Δ” symbol. In HCMVdE::Kan (line 3), the 1-kbp fragment from the kanamycin resistance gene that replaces the HCMV MIEP enhancer is represented by a cross-hatched box. A new EcoRI restriction site was introduced at the location of the enhancer deletion (in HCMVdE) or the stuffer insertion (in HCMVdE::Kan) to facilitate the characterization of the mutant genomes by an EcoRI restriction digestion (data not shown). Sizes of the natural and new EcoRI J DNA fragments for each recombinant BAC are indicated. The illustration is not drawn to scale. (B) Transfection of HCMV enhancerless genomes in cultured fibroblasts. MRC-5 cells were cotransfected by the calcium phosphate precipitation technique with 2 μg of either HCMV (D to F), HCMVdE (G to I and M to O), or HCMVdE::Kan (J to L and P to R) BAC DNAs together with 1 μg of a vector expressing the tegument protein pp71 (2), and when indicated with 1 μg of the HCMV IE1 and IE2 expression vector pSVH (29). GFP expression was detected by fluorescence microscopy at the indicated days after infection (dpi). Magnification, ×20. (C) Growth kinetics of HCMVdE-rev. MRC-5 cells were infected at a multiplicity of infection (MOI) of 0.025 with HCMV or HCMVdE-rev, a revertant of HCMVdE in which sequences from −52 to −667 of the MIEP were reintroduced in the HCMVdE genome by the ET BAC mutagenesis method. At the indicated days postinfection supernatants from the infected cultures were harvested, and titers were determined by standard plaque assays on MRC-5 cells. Each datum point represents the average and standard deviation from three separate cultures. The dashed line represents the limit of detection.
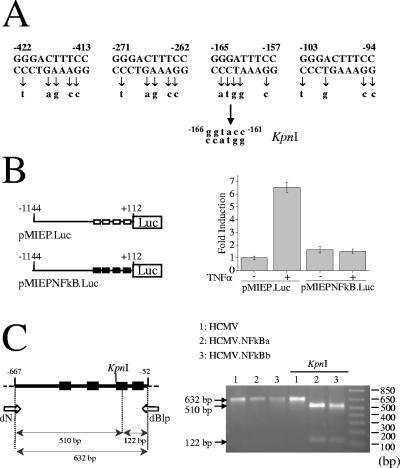
Construction of recombinant HCMVs with point mutations in the NF-κB binding sites of the enhancer region. (A) The sequence and location of the four NF-κB binding sites within the HCMV MIEP are shown, and the point mutations introduced in each specific element are indicated below the wild-type sequence. Mutation of the NF-κB recognition site at position −157 to −165 generates a KpnI restriction site at position −161 to −166, which is shown. Coordinates refer to the HCMV ie1/ie2 transcription start site. (B) The structures of the luciferase (Luc) reporter constructs pMIEP.Luc and pMIEPNFkB.Luc containing MIEP sequences from −1144 to +112 (relative to the HCMV ie1/ie2 transcription start site) without (□) and with (▪) the NF-κB binding sites disrupted, respectively, are shown. 5 × 105 U937 cells were electroporated with 2 μg of pMIEP.Luc or pMIEPNFkB.Luc, along with 0.6 μg of the internal control plasmid pRL-TK, and cultured for 46 h before the luciferase activity was assayed. Six hours before harvesting, cultures were treated with TNF-α (10 ng/ml; +) or vehicle (phosphate-buffered saline; −). The results are presented as the fold induction, taking “1” as the activity presented by the pMIEP.Luc in the absence of TNF-α. The values shown represent the average ± the standard deviation (bars) of four determinations. (C) Schematic diagram of the HCMV MIEP enhancer region, with the location of the four NF-κB binding sites marked by solid boxes. The KpnI restriction site at position −161 to −166 introduced when the NF-κB binding site located between positions −157 and −165 is mutated is indicated, as well as the sizes of the expected KpnI fragments derived from the PCR-amplified enhancer fragment with primers dN and dBlp (3) (flanking the enhancer and indicated with white arrows). Coordinates refer to the HCMV ie1/ie2 transcription start site. To confirm the correct mutagenesis of the NF-κB binding sites, enhancer sequences were amplified from stocks of HCMV (lanes 1 and 4), HCMV.NFkBa (lanes 2 and 5), and HCMV.NFkBb (lanes 3 and 6) by PCR. Marked by arrows are the amplified products before (lanes 1 to 3) or after (lanes 4 to 6) digestion with KpnI resolved by gel electrophoresis. Size markers are shown at the right margin.
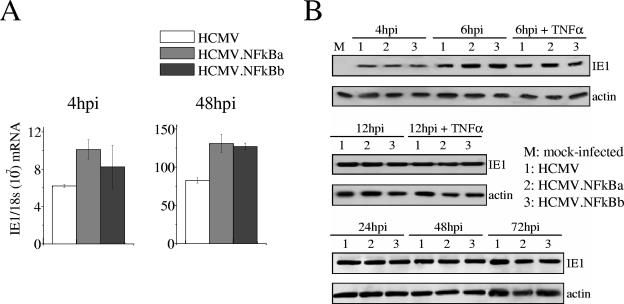
Analysis of ie1 gene expression in HCMV.NFkB-infected cells. (A) Real-time PCR analysis of ie1 RNA expression in cells infected with HCMV.NFkB. HEL fibroblasts were infected at an MOI of 0.01 with HCMV, HCMV.NFkBa, or HCMV.NFkBb and harvested at the time points after infection indicated for isolation of RNA and subsequent analysis by real-time PCR using primers within exon 4 of the HCMV ie1 gene as previously described (3). The results are presented as the relative amount of ie1 mRNA normalized to 18S rRNA, and error bars represent the standard errors of the means. (B) Expression kinetics of the IE1 protein by HCMV.NFkB mutants. HEL fibroblasts were mock infected or infected at an MOI of 0.6 (for the 4-, 6-, and 12-h time points) or 0.1 (for the 24, 48, and 72 h time points). Where indicated, cells were treated with 10 ng of TNF-α/ml 2 h before infection, during, and immediately after the adsorption period. At the indicated time (in hours) postinfection (hpi), samples were lysed, subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis on 7% gels, transferred to nitrocellulose as previously described (1), and probed with an HCMV IE1 specific monoclonal antibody (MAB810; Chemicon, Temecula, CA). As an internal control, actin immunodetection was performed with a monoclonal antibody (A2066; Sigma, St. Louis, MO).
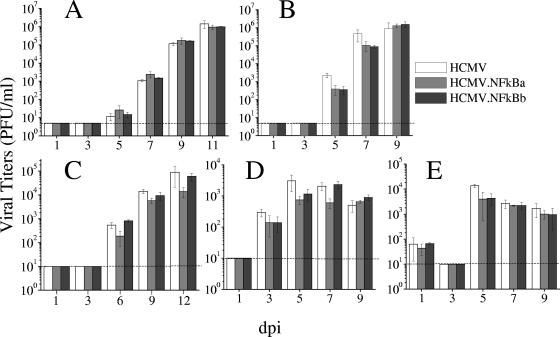
Growth kinetics of HCMV.NFkB mutants. HEL (A), MRC-5 (B), RPE (C), U373 (D), and NT2/D1 cells (E; differentiated for 5 days with 10−5 M retinoic acid) were infected at an MOI of 0.025 (HEL and MRC-5) or 1 (RPE, U373, and NT2/D1, resulting in ca. 10 to 25% of GFP-positive cells in the culture at 48 hpi) with HCMV, HCMV.NFkBa, or HCMV.NFkBb. At the indicated time in days postinfection, the amount of extracellular (HEL, MRC-5, and RPE) or cell associated (U373 and NT2/D1) infectious virus present in the cultures was determined by plaque titration assays on MRC-5 cells. Each datum point represents the average and standard deviation from three separate cultures. Dashed lines represent the limits of detection.
Similar articles
-
The Elk-1 and serum response factor binding sites in the major immediate-early promoter of human cytomegalovirus are required for efficient viral replication in quiescent cells and compensate for inactivation of the NF-kappaB sites in proliferating cells.J Virol. 2010 May;84(9):4481-93. doi: 10.1128/JVI.02141-09. Epub 2010 Feb 10. J Virol. 2010. PMID: 20147408 Free PMC article.
-
Activation of the virus-induced IKK/NF-kappaB signalling axis is critical for the replication of human cytomegalovirus in quiescent cells.Cell Microbiol. 2007 Aug;9(8):2040-54. doi: 10.1111/j.1462-5822.2007.00936.x. Epub 2007 Apr 5. Cell Microbiol. 2007. PMID: 17419715
-
Two gamma interferon-activated site-like elements in the human cytomegalovirus major immediate-early promoter/enhancer are important for viral replication.J Virol. 2005 Apr;79(8):5035-46. doi: 10.1128/JVI.79.8.5035-5046.2005. J Virol. 2005. PMID: 15795289 Free PMC article.
-
Immediate-early transcription regulation of human cytomegalovirus.Curr Top Microbiol Immunol. 1990;154:3-19. doi: 10.1007/978-3-642-74980-3_1. Curr Top Microbiol Immunol. 1990. PMID: 2161324 Review. No abstract available.
-
Role of the cytomegalovirus major immediate early enhancer in acute infection and reactivation from latency.Med Microbiol Immunol. 2008 Jun;197(2):223-31. doi: 10.1007/s00430-007-0069-7. Epub 2007 Dec 19. Med Microbiol Immunol. 2008. PMID: 18097687 Review.
Cited by
-
Phorbol ester-induced human cytomegalovirus major immediate-early (MIE) enhancer activation through PKC-delta, CREB, and NF-kappaB desilences MIE gene expression in quiescently infected human pluripotent NTera2 cells.J Virol. 2010 Sep;84(17):8495-508. doi: 10.1128/JVI.00416-10. Epub 2010 May 26. J Virol. 2010. PMID: 20504934 Free PMC article.
-
New cell-signaling pathways for controlling cytomegalovirus replication.Am J Transplant. 2014 Jun;14(6):1249-58. doi: 10.1111/ajt.12725. Epub 2014 May 19. Am J Transplant. 2014. PMID: 24839861 Free PMC article. Review.
-
cGAS-STING-TBK1 Signaling Promotes Valproic Acid-Responsive Human Cytomegalovirus Immediate-Early Transcription during Infection of Incompletely Differentiated Myeloid Cells.Viruses. 2024 May 30;16(6):877. doi: 10.3390/v16060877. Viruses. 2024. PMID: 38932169 Free PMC article.
-
Proinflammatory cytokine-induced tight junction remodeling through dynamic self-assembly of claudins.Mol Biol Cell. 2014 Sep 15;25(18):2710-9. doi: 10.1091/mbc.E14-02-0773. Epub 2014 Jul 16. Mol Biol Cell. 2014. PMID: 25031428 Free PMC article.
-
NFκB and Cyclic AMP Response Element Sites Mediate the Valproic Acid and UL138 Responsiveness of the Human Cytomegalovirus Major Immediate Early Enhancer and Promoter.J Virol. 2023 Mar 30;97(3):e0002923. doi: 10.1128/jvi.00029-23. Epub 2023 Mar 1. J Virol. 2023. PMID: 36856444 Free PMC article.
References
-
- Benedict, C. A., T. A. Banks, L. Senderowicz, M. Ko, W. J. Britt, A. Angulo, P. Ghazal, and C. F. Ware. 2001. Lymphotoxins and cytomegalovirus cooperatively induce interferon-beta, establishing host-virus detente. Immunity 15:617-626. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
