

Twin gradients in APOBEC3 edited HIV-1 DNA reflect the dynamics of lentiviral replication
- PMID: 16963778
- PMCID: PMC1635257
- DOI: 10.1093/nar/gkl555
Twin gradients in APOBEC3 edited HIV-1 DNA reflect the dynamics of lentiviral replication
Abstract
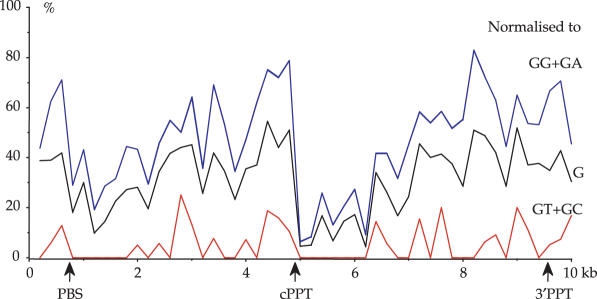
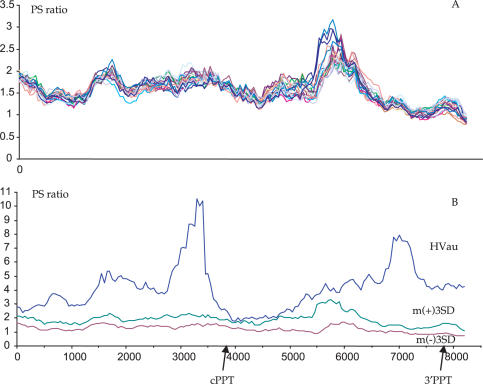
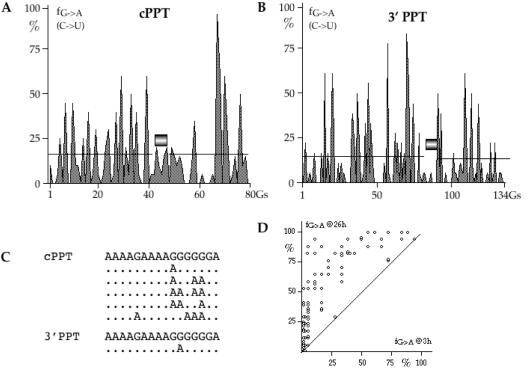
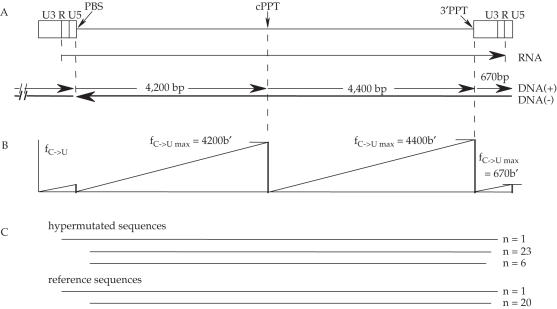
The human immunodeficiency virus (HIV) Vif protein blocks incorporation of two host cell cytidine deaminases, APOBEC3F and 3G, into the budding virion. Not surprisingly, on a vif background nascent minus strand DNA can be extensively edited leaving multiple uracil residues. Editing occurs preferentially in the context of TC (GA on the plus strand) and CC (GG) depending on the enzyme. To explore the distribution of APOBEC3F and -3G editing across the genome, a product/substrate ratio (AA + AG)/(GA + GG) was computed for a series of 30 edited genomes present in the data bases. Two highly polarized gradients were noted each with maxima just 5' to the central polypurine tract (cPPT) and LTR proximal polypurine tract (3'PPT). The gradients are in remarkable agreement with the time the minus strand DNA remains single stranded. In vitro analyses of APOBEC3G deamination of nascent cDNA spanning the two PPTs showed no pronounced dependence on the PPT RNA:DNA heteroduplex ruling out the competing hypothesis of a PPT orientation effect. The degree of hypermutation varied smoothly among genomes indicating that the number of APOBEC3 molecules packaged varied considerably.
Figures


Similar articles
-
Polymorphisms of the cytidine deaminase APOBEC3F have different HIV-1 restriction efficiencies.Virology. 2019 Jan 15;527:21-31. doi: 10.1016/j.virol.2018.11.004. Epub 2018 Nov 15. Virology. 2019. PMID: 30448640
-
Extensive editing of a small fraction of human T-cell leukemia virus type 1 genomes by four APOBEC3 cytidine deaminases.J Gen Virol. 2005 Sep;86(Pt 9):2489-2494. doi: 10.1099/vir.0.80973-0. J Gen Virol. 2005. PMID: 16099907
-
APOBEC3F properties and hypermutation preferences indicate activity against HIV-1 in vivo.Curr Biol. 2004 Aug 10;14(15):1385-91. doi: 10.1016/j.cub.2004.06.050. Curr Biol. 2004. PMID: 15296757
-
Hepatitis B: modern concepts in pathogenesis--APOBEC3 cytidine deaminases as effectors in innate immunity against the hepatitis B virus.Curr Opin Infect Dis. 2008 Jun;21(3):298-303. doi: 10.1097/QCO.0b013e3282fe1bb2. Curr Opin Infect Dis. 2008. PMID: 18448976 Review.
-
Cytidine deaminases as a weapon against retroviruses and a new target for antiviral therapy.Mini Rev Med Chem. 2008 Mar;8(3):231-8. doi: 10.2174/138955708783744047. Mini Rev Med Chem. 2008. PMID: 18336343 Review.
Cited by
-
In search of a function for the most frequent naturally-occurring length polymorphism (MFNLP) of the HIV-1 LTR: retaining functional coupling, of Nef and RBF-2, at RBEIII?Int J Biol Sci. 2007 Jun 11;3(5):318-27. doi: 10.7150/ijbs.3.318. Int J Biol Sci. 2007. PMID: 17589566 Free PMC article. Review.
-
Evidence for restriction of ancient primate gammaretroviruses by APOBEC3 but not TRIM5alpha proteins.PLoS Pathog. 2008 Oct;4(10):e1000181. doi: 10.1371/journal.ppat.1000181. Epub 2008 Oct 17. PLoS Pathog. 2008. PMID: 18927623 Free PMC article.
-
Insights into the motif preference of APOBEC3 enzymes.PLoS One. 2014 Jan 31;9(1):e87679. doi: 10.1371/journal.pone.0087679. eCollection 2014. PLoS One. 2014. PMID: 24498164 Free PMC article.
-
Human immunodeficiency virus (HIV) type 1 proviral hypermutation correlates with CD4 count in HIV-infected women from Kenya.J Virol. 2008 Aug;82(16):8172-82. doi: 10.1128/JVI.01115-08. Epub 2008 Jun 11. J Virol. 2008. PMID: 18550667 Free PMC article.
-
A model for oligomeric regulation of APOBEC3G cytosine deaminase-dependent restriction of HIV.J Biol Chem. 2008 May 16;283(20):13780-91. doi: 10.1074/jbc.M801004200. Epub 2008 Mar 24. J Biol Chem. 2008. PMID: 18362149 Free PMC article.
References
-
- Beale R.C., Petersen-Mahrt S.K., Watt I.N., Harris R.S., Rada C., Neuberger M.S. Comparison of the differential context-dependence of DNA deamination by APOBEC enzymes: correlation with mutation spectra in vivo. J. Mol. Biol. 2004;337:585–596. - PubMed
-
- Bishop K.N., Holmes R.K., Sheehy A.M., Davidson N.O., Cho S.J., Malim M.H. Cytidine deamination of retroviral DNA by diverse APOBEC proteins. Curr. Biol. 2004;14:1392–1396. - PubMed
-
- Harris R.S., Bishop K.N., Sheehy A.M., Craig H.M., Petersen-Mahrt S.K., Watt I.N., Neuberger M.S., Malim M.H. DNA deamination mediates innate immunity to retroviral infection. Cell. 2003;113:803–809. - PubMed
-
- Lecossier D., Bouchonnet F., Clavel F., Hance A.J. Hypermutation of HIV-1 DNA in the absence of the Vif protein. Science. 2003;300:1112. - PubMed
-
- Liddament M.T., Brown W.L., Schumacher A.J., Harris R.S. APOBEC3F properties and hypermutation preferences indicate activity against HIV-1 in vivo. Curr. Biol. 2004;14:1385–1391. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
