

Generation and characterization of B7-H4/B7S1/B7x-deficient mice
- PMID: 16914726
- PMCID: PMC1592821
- DOI: 10.1128/MCB.00755-06
Generation and characterization of B7-H4/B7S1/B7x-deficient mice
Abstract
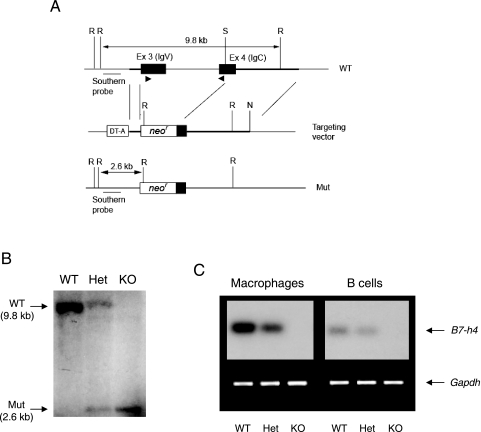
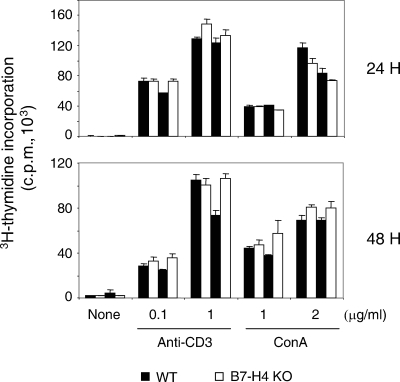
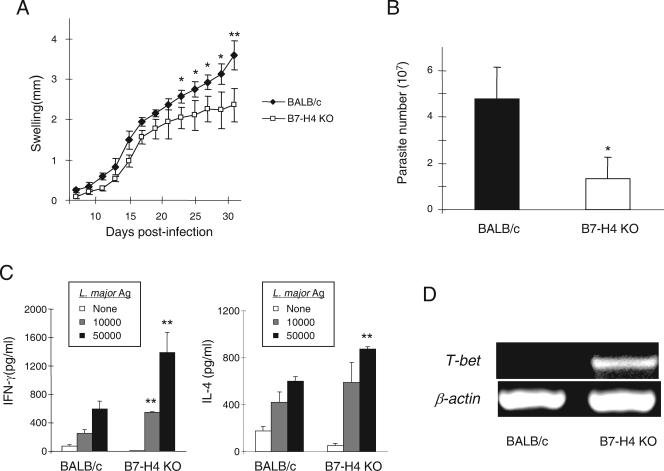
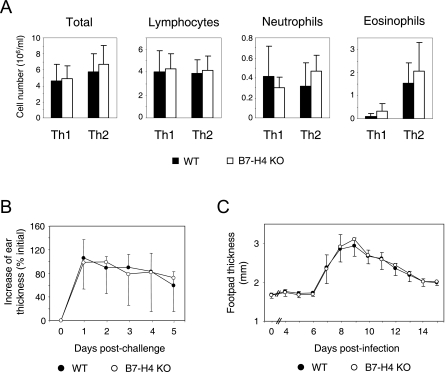
Members of the B7 family of cosignaling molecules regulate T-cell proliferation and effector functions by engaging cognate receptors on T cells. In vitro and in vivo blockade experiments indicated that B7-H4 (also known as B7S1 or B7x) inhibits proliferation, cytokine production, and cytotoxicity of T cells. B7-H4 binds to an unknown receptor(s) that is expressed on activated T cells. However, whether B7-H4 plays nonredundant immune regulatory roles in vivo has not been tested. We generated B7-H4-deficient mice to investigate the roles of B7-H4 during various immune reactions. Consistent with its inhibitory function in vitro, B7-H4-deficient mice mounted mildly augmented T-helper 1 (Th1) responses and displayed slightly lowered parasite burdens upon Leishmania major infection compared to the wild-type mice. However, the lack of B7-H4 did not affect hypersensitive inflammatory responses in the airway or skin that are induced by either Th1 or Th2 cells. Likewise, B7-H4-deficient mice developed normal cytotoxic T-lymphocyte reactions against viral infection. Thus, B7-H4 plays a negative regulatory role in vivo but the impact of B7-H4 deficiency is minimal. These results suggest that B7-H4 is one of multiple negative cosignaling molecules that collectively provide a fine-tuning mechanism for T-cell-mediated immune responses.
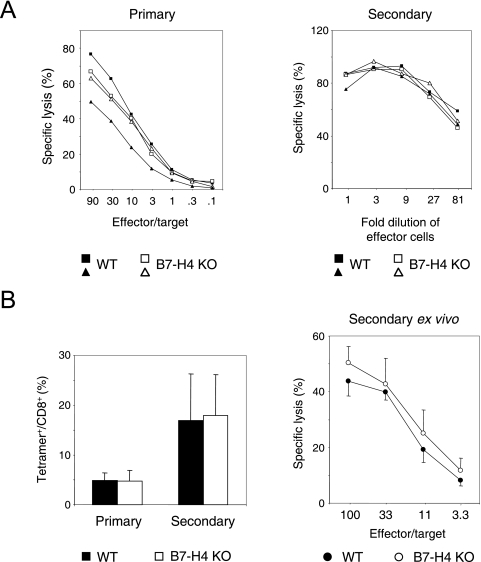
Figures
Similar articles
-
The B7 family member B7-H3 preferentially down-regulates T helper type 1-mediated immune responses.Nat Immunol. 2003 Sep;4(9):899-906. doi: 10.1038/ni967. Epub 2003 Aug 17. Nat Immunol. 2003. PMID: 12925852
-
Studies using antigen-presenting cells lacking expression of both B7-1 (CD80) and B7-2 (CD86) show distinct requirements for B7 molecules during priming versus restimulation of Th2 but not Th1 cytokine production.J Immunol. 1998 Sep 15;161(6):2762-71. J Immunol. 1998. PMID: 9743334
-
Possible involvement of soluble B7-H4 in T cell-mediated inflammatory immune responses.Biochem Biophys Res Commun. 2009 Nov 13;389(2):349-53. doi: 10.1016/j.bbrc.2009.08.144. Epub 2009 Aug 31. Biochem Biophys Res Commun. 2009. PMID: 19723502
-
Potential targeting of B7-H4 for the treatment of cancer.Immunol Rev. 2017 Mar;276(1):40-51. doi: 10.1111/imr.12530. Immunol Rev. 2017. PMID: 28258701 Free PMC article. Review.
-
B7x-from bench to bedside.ESMO Open. 2019 Sep 8;4(5):e000554. doi: 10.1136/esmoopen-2019-000554. eCollection 2019. ESMO Open. 2019. PMID: 31555486 Free PMC article. Review.
Cited by
-
The B7x Immune Checkpoint Pathway: From Discovery to Clinical Trial.Trends Pharmacol Sci. 2019 Nov;40(11):883-896. doi: 10.1016/j.tips.2019.09.008. Epub 2019 Oct 31. Trends Pharmacol Sci. 2019. PMID: 31677920 Free PMC article. Review.
-
Costimulatory and coinhibitory receptors in anti-tumor immunity.Immunol Rev. 2009 May;229(1):126-44. doi: 10.1111/j.1600-065X.2009.00771.x. Immunol Rev. 2009. PMID: 19426219 Free PMC article. Review.
-
The nature of activatory and tolerogenic dendritic cell-derived signal II.Front Immunol. 2013 Feb 28;4:53. doi: 10.3389/fimmu.2013.00053. eCollection 2013. Front Immunol. 2013. PMID: 23450201 Free PMC article.
-
B7-H4-deficient mice display augmented neutrophil-mediated innate immunity.Blood. 2009 Feb 19;113(8):1759-67. doi: 10.1182/blood-2008-01-133223. Epub 2008 Dec 24. Blood. 2009. PMID: 19109567 Free PMC article.
-
Tumor Regression and Delayed Onset Toxicity Following B7-H4 CAR T Cell Therapy.Mol Ther. 2016 Nov;24(11):1987-1999. doi: 10.1038/mt.2016.149. Epub 2016 Jul 21. Mol Ther. 2016. PMID: 27439899 Free PMC article.
References
-
- Bertram, E. M., P. Lau, and T. H. Watts. 2002. Temporal segregation of 4-1BB versus CD28-mediated costimulation: 4-1BB ligand influences T cell numbers late in the primary response and regulates the size of the T cell memory response following influenza infection. J. Immunol. 168:3777-3785. - PubMed
-
- Bretscher, P., and M. Cohn. 1970. A theory of self-nonself discrimination. Science 169:1042-1049. - PubMed
-
- Chapoval, A. I., J. Ni, J. S. Lau, R. A. Wilcox, D. B. Flies, D. Liu, H. Dong, G. L. Sica, G. Zhu, K. Tamada, and L. Chen. 2001. B7-H3: a costimulatory molecule for T cell activation and IFN-γ production. Nat. Immunol. 2:269-274. - PubMed
-
- Choi, I. H., G. Zhu, G. L. Sica, S. E. Strome, J. C. Cheville, J. S. Lau, Y. Zhu, D. B. Flies, K. Tamada, and L. Chen. 2003. Genomic organization and expression analysis of B7-H4, an immune inhibitory molecule of the B7 family. J. Immunol. 171:4650-4654. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials