

Functional cooperation between FACT and MCM helicase facilitates initiation of chromatin DNA replication
- PMID: 16902406
- PMCID: PMC1560368
- DOI: 10.1038/sj.emboj.7601271
Functional cooperation between FACT and MCM helicase facilitates initiation of chromatin DNA replication
Abstract
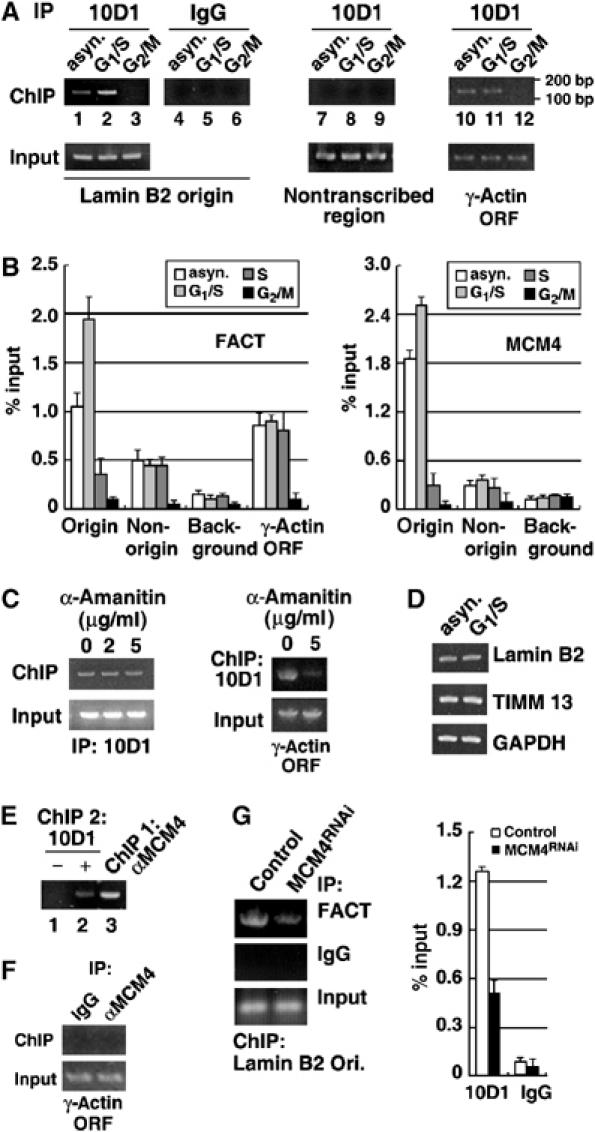
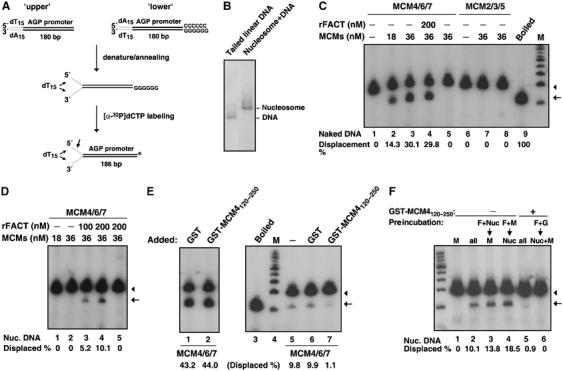
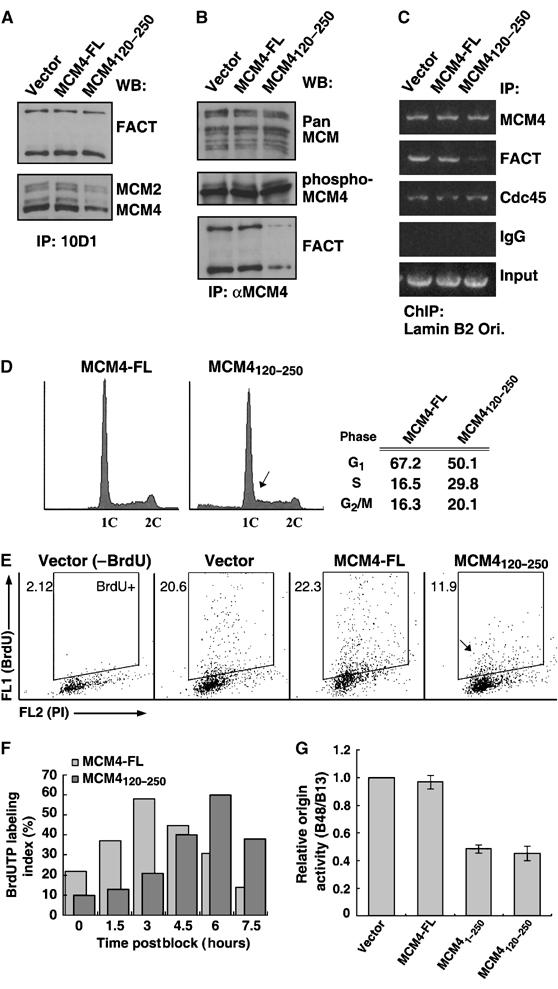
Chromatin is suppressive in nature to cellular enzymes that metabolize DNA, mainly due to the inherent inaccessibility of the DNA template. Despite extensive understanding of the involvement of chromatin-modifying factors in transcription, roles of related activities in DNA replication remain largely elusive. Here, we show that the heterodimeric transcriptional elongation factor FACT (facilitates chromatin transcription) is functionally linked to DNA synthesis. Its involvement in DNA replication is partly mediated by the stable association with the replicative helicase complex, MCM, and further by the coexistence with MCM on replication origin. Furthermore, relying on its nucleosome-reorganizing activity, FACT can facilitate chromatin unwinding by the MCM complex, which is otherwise inert on the nucleosomal template. As a consequence, the physical and functional interaction between FACT and MCM is an important determinant in the proper initiation of DNA replication and S phase in vivo. Together, our findings identify FACT as an integral and conserved component of the endogenous replication machinery, and support a model in which the concerted action of helicase and chromatin-modifying activities promotes chromosome replication.
Figures

Similar articles
-
Keys to open chromatin for transcription activation: FACT and Asf1.Mol Cell. 2009 May 14;34(4):397-9. doi: 10.1016/j.molcel.2009.05.004. Mol Cell. 2009. PMID: 19481517
-
Functional cooperation between FACT and MCM is coordinated with cell cycle and differential complex formation.J Biomed Sci. 2010 Feb 16;17(1):11. doi: 10.1186/1423-0127-17-11. J Biomed Sci. 2010. PMID: 20156367 Free PMC article.
-
The histone chaperone facilitates chromatin transcription (FACT) protein maintains normal replication fork rates.J Biol Chem. 2011 Sep 2;286(35):30504-30512. doi: 10.1074/jbc.M111.264721. Epub 2011 Jul 7. J Biol Chem. 2011. PMID: 21757688 Free PMC article.
-
FACT and the reorganized nucleosome.Mol Biosyst. 2008 Nov;4(11):1085-93. doi: 10.1039/b812136b. Epub 2008 Sep 19. Mol Biosyst. 2008. PMID: 18931784 Review.
-
Control of DNA replication: regulation and activation of eukaryotic replicative helicase, MCM.IUBMB Life. 2005 Apr-May;57(4-5):323-35. doi: 10.1080/15216540500092419. IUBMB Life. 2005. PMID: 16036617 Review.
Cited by
-
Regulation of Unperturbed DNA Replication by Ubiquitylation.Genes (Basel). 2015 Jun 25;6(3):451-68. doi: 10.3390/genes6030451. Genes (Basel). 2015. PMID: 26121093 Free PMC article. Review.
-
The effects of oligomerization on Saccharomyces cerevisiae Mcm4/6/7 function.BMC Biochem. 2010 Sep 22;11:37. doi: 10.1186/1471-2091-11-37. BMC Biochem. 2010. PMID: 20860810 Free PMC article.
-
Identification of Elg1 interaction partners and effects on post-replication chromatin re-formation.PLoS Genet. 2018 Nov 12;14(11):e1007783. doi: 10.1371/journal.pgen.1007783. eCollection 2018 Nov. PLoS Genet. 2018. PMID: 30418970 Free PMC article.
-
The Histone Chaperone FACT Coordinates H2A.X-Dependent Signaling and Repair of DNA Damage.Mol Cell. 2018 Dec 6;72(5):888-901.e7. doi: 10.1016/j.molcel.2018.09.010. Epub 2018 Oct 18. Mol Cell. 2018. PMID: 30344095 Free PMC article.
-
Cyclin-dependent kinase-associated proteins Cks1 and Cks2 are essential during early embryogenesis and for cell cycle progression in somatic cells.Mol Cell Biol. 2008 Sep;28(18):5698-709. doi: 10.1128/MCB.01833-07. Epub 2008 Jul 14. Mol Cell Biol. 2008. PMID: 18625720 Free PMC article.
References
-
- Abdurashidova G, Deganuto M, Klima R, Riva S, Biamonti G, Giacca M, Falaschi A (2000) Start sites of bidirectional DNA synthesis at the human lamin B2 origin. Science 287: 2023–2026 - PubMed
-
- Aggarwal BD, Calvi BR (2004) Chromatin regulates origin activity in Drosophila follicle cells. Nature 430: 372–376 - PubMed
-
- Aparicio OM, Weinstein DM, Bell SP (1997) Components and dynamics of DNA replication complexes in S. cerevisiae: redistribution of MCM proteins and Cdc45p during S phase. Cell 91: 59–69 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
