

Independent contributions of polyomavirus middle T and small T to the regulation of early and late gene expression and DNA replication
- PMID: 16840310
- PMCID: PMC1563708
- DOI: 10.1128/JVI.00679-06
Independent contributions of polyomavirus middle T and small T to the regulation of early and late gene expression and DNA replication
Abstract
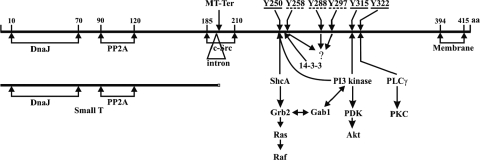
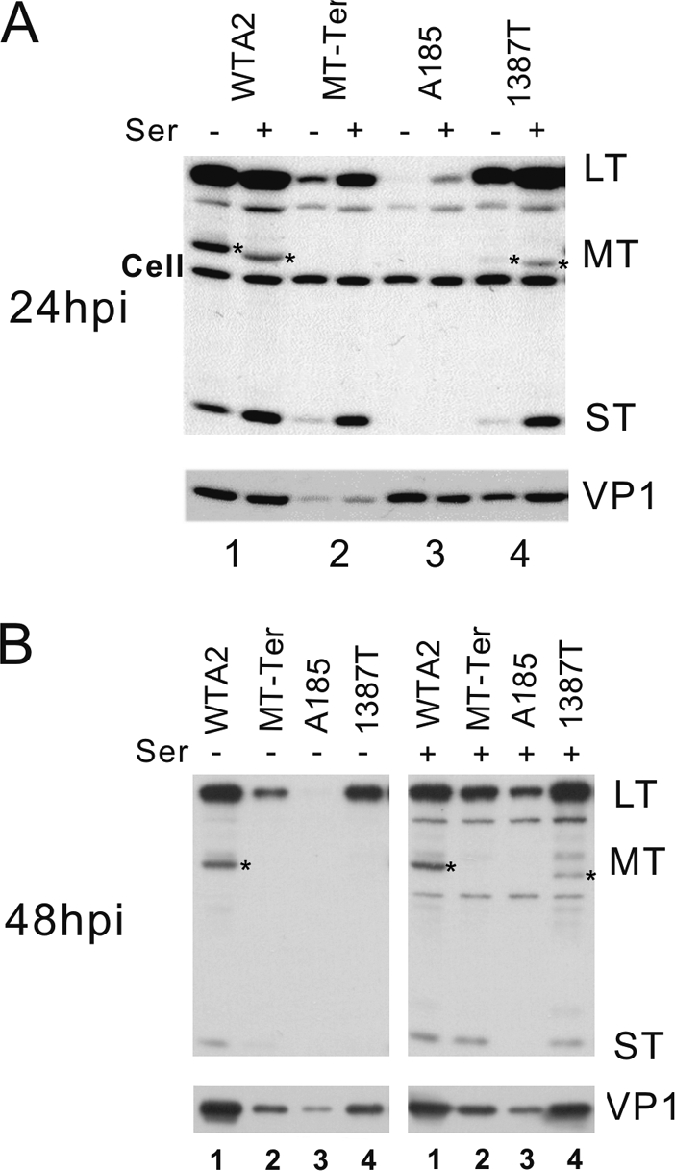
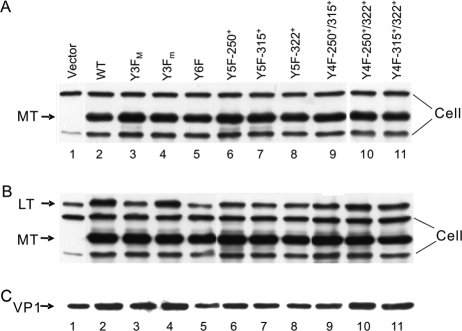
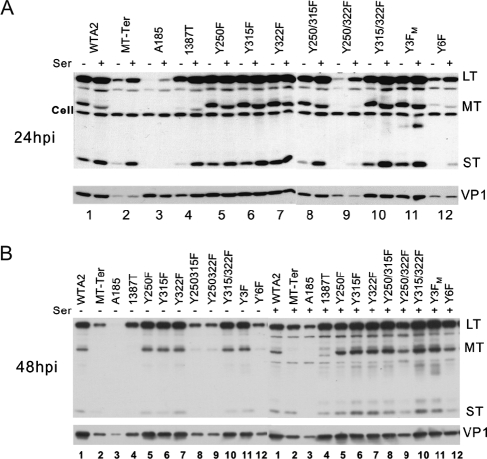
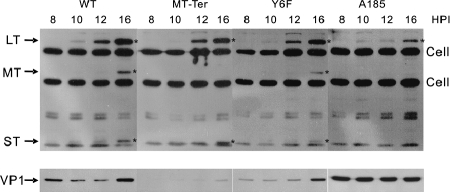
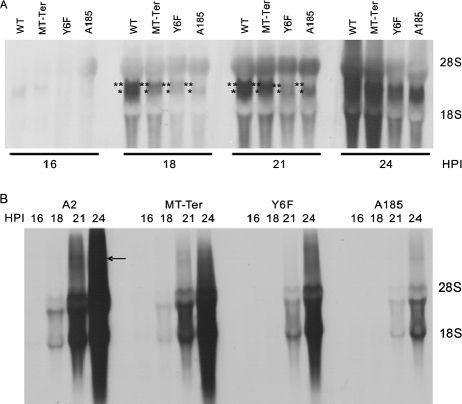
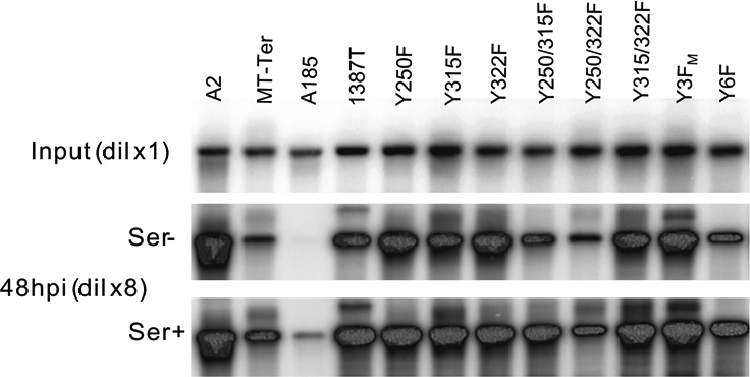
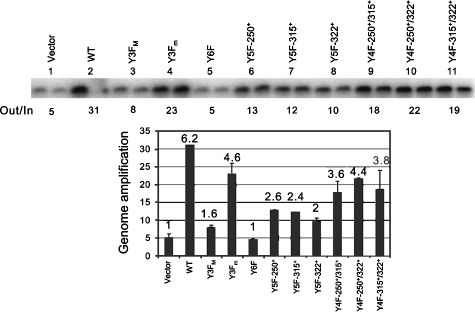
We previously showed that murine polyomavirus mutants that lack both middle T (MT) and small T (ST) functions have a severe pleiotropic defect in early and late viral gene expression as well as genome amplification. The respective contribution of MT and ST to this phenotype was unclear. This work separates the roles of MT and ST in both permissive mouse cells and nonpermissive rat cells. It demonstrates for the first time a role for both proteins. To gain insight into the signaling pathways that might be required, we focused on MT and its mutants. The results show that each of the major MT signaling connections, Shc, phosphatidylinositol 3'-kinase, and phospholipase C gamma1, could contribute in an additive way. Unexpectedly, a mutant lacking all these connections because the three major tyrosines had been converted to phenylalanine retained some activity. A mutant in which all six MT C-terminal tyrosines had been mutated was inactive. This suggests a novel signaling pathway for MT that uses the minor tyrosines. What is common to ST and the individual MT signaling pathways is the ability to signal to the polyomavirus enhancer, in particular to the crucial AP-1 and PEA3/ets binding sites. This connection explains the pleiotropy of MT and ST effects on transcription and DNA replication.
Figures
Similar articles
-
Lessons from polyoma middle T antigen on signaling and transformation: A DNA tumor virus contribution to the war on cancer.Virology. 2009 Feb 20;384(2):304-16. doi: 10.1016/j.virol.2008.09.042. Epub 2008 Nov 20. Virology. 2009. PMID: 19022468 Free PMC article. Review.
-
A Transformation-Defective Polyomavirus Middle T Antigen with a Novel Defect in PI3 Kinase Signaling.J Virol. 2017 Jan 3;91(2):e01774-16. doi: 10.1128/JVI.01774-16. Print 2017 Jan 15. J Virol. 2017. PMID: 27852846 Free PMC article.
-
Signaling from polyomavirus middle T and small T defines different roles for protein phosphatase 2A.Mol Cell Biol. 1998 Dec;18(12):7556-64. doi: 10.1128/MCB.18.12.7556. Mol Cell Biol. 1998. PMID: 9819441 Free PMC article.
-
Transformation by Polyomavirus Middle T Antigen Involves a Unique Bimodal Interaction with the Hippo Effector YAP.J Virol. 2016 Jul 27;90(16):7032-7045. doi: 10.1128/JVI.00417-16. Print 2016 Aug 15. J Virol. 2016. PMID: 27194756 Free PMC article.
-
Natural biology of polyomavirus middle T antigen.Microbiol Mol Biol Rev. 2001 Jun;65(2):288-318 ; second and third pages, table of contents. doi: 10.1128/MMBR.65.2.288-318.2001. Microbiol Mol Biol Rev. 2001. PMID: 11381103 Free PMC article. Review.
Cited by
-
Activation of DNA damage repair pathways by murine polyomavirus.Virology. 2016 Oct;497:346-356. doi: 10.1016/j.virol.2016.07.028. Epub 2016 Aug 16. Virology. 2016. PMID: 27529739 Free PMC article.
-
The p110alpha isoform of phosphatidylinositol 3-kinase is essential for polyomavirus middle T antigen-mediated transformation.J Virol. 2007 Jul;81(13):7069-76. doi: 10.1128/JVI.00115-07. Epub 2007 Apr 18. J Virol. 2007. PMID: 17442716 Free PMC article.
-
Lessons from polyoma middle T antigen on signaling and transformation: A DNA tumor virus contribution to the war on cancer.Virology. 2009 Feb 20;384(2):304-16. doi: 10.1016/j.virol.2008.09.042. Epub 2008 Nov 20. Virology. 2009. PMID: 19022468 Free PMC article. Review.
-
Lessons in signaling and tumorigenesis from polyomavirus middle T antigen.Microbiol Mol Biol Rev. 2009 Sep;73(3):542-63, Table of Contents. doi: 10.1128/MMBR.00009-09. Microbiol Mol Biol Rev. 2009. PMID: 19721090 Free PMC article. Review.
-
Cellular transformation by Simian Virus 40 and Murine Polyoma Virus T antigens.Semin Cancer Biol. 2009 Aug;19(4):218-28. doi: 10.1016/j.semcancer.2009.03.002. Epub 2009 Mar 31. Semin Cancer Biol. 2009. PMID: 19505649 Free PMC article. Review.
References
-
- Besser, D., M. Urich, M. Sakaue, A. Messerschmitt, K. Ballmer-Hofer, and Y. Nagamine. 1995. Urokinase-type plasminogen activator gene regulation by polyomavirus middle-T antigen. Oncogene 11:2383-2391. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
