

CD1d mediates T-cell-dependent resistance to secondary infection with encephalomyocarditis virus (EMCV) in vitro and immune response to EMCV infection in vivo
- PMID: 16809320
- PMCID: PMC1489038
- DOI: 10.1128/JVI.02745-05
CD1d mediates T-cell-dependent resistance to secondary infection with encephalomyocarditis virus (EMCV) in vitro and immune response to EMCV infection in vivo
Abstract
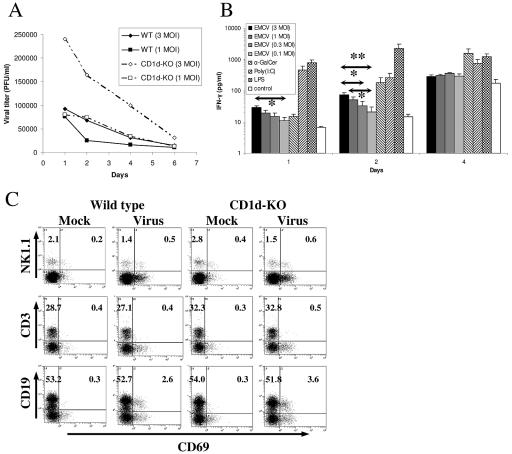
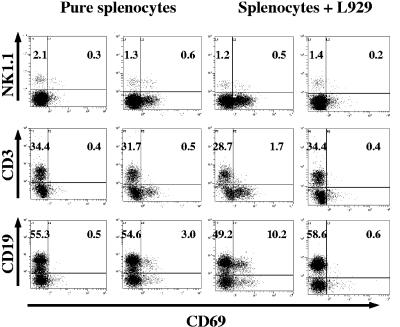
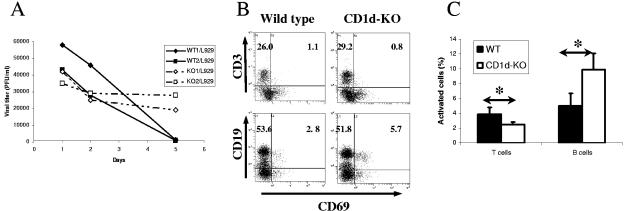
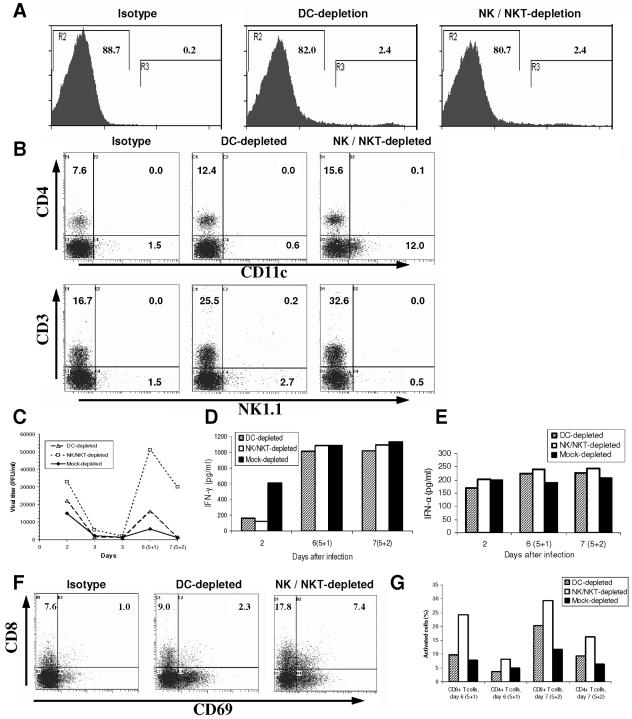
The innate and adaptive immune responses have evolved distinct strategies for controlling different viral pathogens. Encephalomyocarditis virus (EMCV) is a picornavirus that can cause paralysis, diabetes, and myocarditis within days of infection. The optimal innate immune response against EMCV in vivo requires CD1d. Interaction of antigen-presenting cell CD1d with distinct natural killer T-cell ("NKT") populations can induce rapid gamma interferon (IFN-gamma) production and NK-cell activation. The T-cell response of CD1d-deficient mice (lacking all NKT cells) against acute EMCV infection was further studied in vitro and in vivo. EMCV persisted at higher levels in CD1d-knockout (KO) splenocyte cultures infected in vitro. Furthermore, optimal resistance to repeat cycles of EMCV infection in vitro was also shown to depend on CD1d. However, this was not reflected in the relative levels of NK-cell activation but rather by the responses of both CD4(+) and CD8(+) T-cell populations. Repeated EMCV infection in vitro induced less IFN-gamma and alpha interferon (IFN-alpha) from CD1d-deficient splenocytes than with the wild type. Furthermore, the level of EMCV replication in wild-type splenocytes was markedly and specifically increased by addition of blocking anti-CD1d antibody. Depletion experiments demonstrated that dendritic cells contributed less than the combination of NK and NKT cells to anti-EMCV responses and that none of these cell types was the main source of IFN-alpha. Finally, EMCV infection in vivo produced higher levels of viremia in CD1d-KO mice than in wild-type animals, coupled with significantly less lymphocyte activation and IFN-alpha production. These results point to the existence of a previously unrecognized mechanism of rapid CD1d-dependent stimulation of the antiviral adaptive cellular immune response.
Figures





Similar articles
-
Innate immune response to encephalomyocarditis virus infection mediated by CD1d.Immunology. 2003 Dec;110(4):519-26. doi: 10.1111/j.1365-2567.2003.01779.x. Immunology. 2003. PMID: 14632651 Free PMC article.
-
Direct CD1d-mediated stimulation of APC IL-12 production and protective immune response to virus infection in vivo.J Immunol. 2010 Jan 1;184(1):268-76. doi: 10.4049/jimmunol.0800924. Epub 2009 Nov 30. J Immunol. 2010. PMID: 19949077 Free PMC article.
-
CD1d-reactive T-cell activation leads to amelioration of disease caused by diabetogenic encephalomyocarditis virus.J Leukoc Biol. 2001 May;69(5):713-8. J Leukoc Biol. 2001. PMID: 11358978
-
Regulation of immune responses by CD1d-restricted natural killer T cells.Immunol Res. 2004;30(2):139-53. doi: 10.1385/IR:30:2:139. Immunol Res. 2004. PMID: 15477656 Review.
-
CD1 antigen presentation and infectious disease.Contrib Microbiol. 2003;10:164-82. doi: 10.1159/000068136. Contrib Microbiol. 2003. PMID: 12530326 Review.
Cited by
-
Herpes Simplex Virus 1 Specifically Targets Human CD1d Antigen Presentation To Enhance Its Pathogenicity.J Virol. 2018 Oct 29;92(22):e01490-18. doi: 10.1128/JVI.01490-18. Print 2018 Nov 15. J Virol. 2018. PMID: 30185591 Free PMC article.
-
Contribution of autophagy to antiviral immunity.FEBS Lett. 2015 Nov 14;589(22):3461-70. doi: 10.1016/j.febslet.2015.07.047. Epub 2015 Aug 20. FEBS Lett. 2015. PMID: 26297829 Free PMC article. Review.
-
Tolerance and Cross-Tolerance following Toll-Like Receptor (TLR)-4 and -9 Activation Are Mediated by IRAK-M and Modulated by IL-7 in Murine Splenocytes.PLoS One. 2015 Jul 28;10(7):e0132921. doi: 10.1371/journal.pone.0132921. eCollection 2015. PLoS One. 2015. PMID: 26218271 Free PMC article.
-
Human cytomegalovirus (HCMV) US2 protein interacts with human CD1d (hCD1d) and down-regulates invariant NKT (iNKT) cell activity.Mol Cells. 2013 Nov;36(5):455-64. doi: 10.1007/s10059-013-0221-8. Epub 2013 Nov 8. Mol Cells. 2013. PMID: 24213674 Free PMC article.
-
The immune response to papillomavirus during infection persistence and regression.Open Virol J. 2012;6:241-8. doi: 10.2174/1874357901206010241. Epub 2012 Dec 28. Open Virol J. 2012. PMID: 23341859 Free PMC article.
References
-
- Baron, J. L., L. Gardiner, S. Nishimura, K. Shinkai, R. Locksley, and D. Ganem. 2002. Activation of a nonclassical NKT cell subset in a transgenic mouse model of hepatitis B virus infection. Immunity 16:583-594. - PubMed
-
- Behar, S. M., and S. Cardell. 2000. Diverse CD1d-restricted T cells: diverse phenotypes, and diverse functions. Semin. Immunol. 12:551-560. - PubMed
-
- Behar, S. M., T. A. Podrebarac, C. J. Roy, C. R. Wang, and M. B. Brenner. 1999. Diverse TCRs recognize murine CD1. J. Immunol. 162:161-167. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
