

Cytosolic chaperonin protects folding intermediates of Gbeta from aggregation by recognizing hydrophobic beta-strands
- PMID: 16717193
- PMCID: PMC1482499
- DOI: 10.1073/pnas.0600195103
Cytosolic chaperonin protects folding intermediates of Gbeta from aggregation by recognizing hydrophobic beta-strands
Abstract
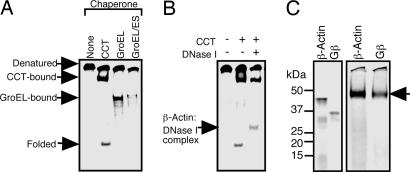
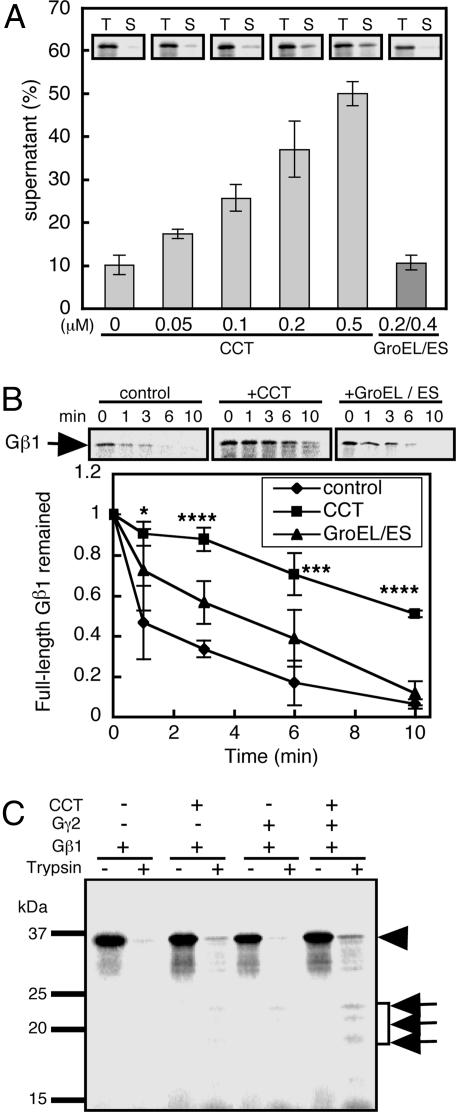
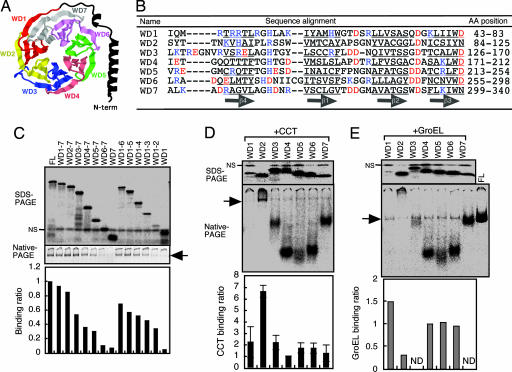
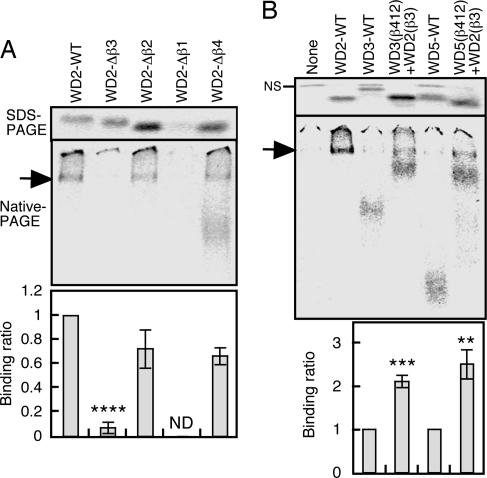
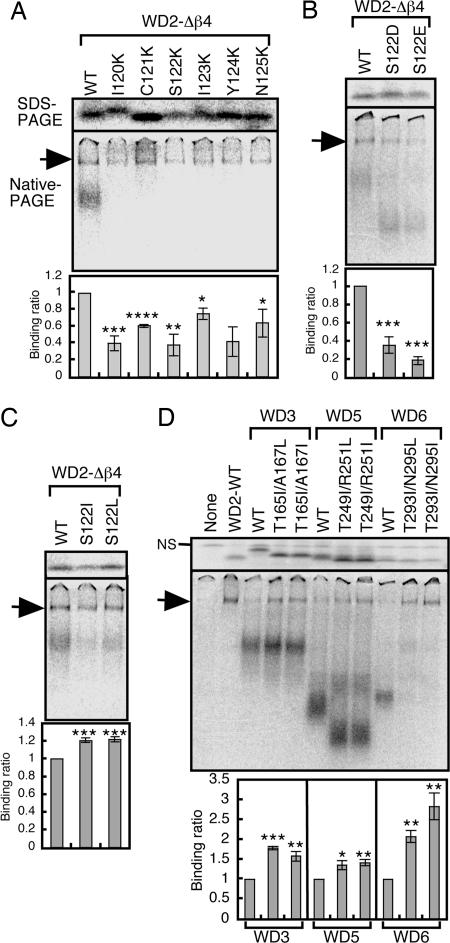
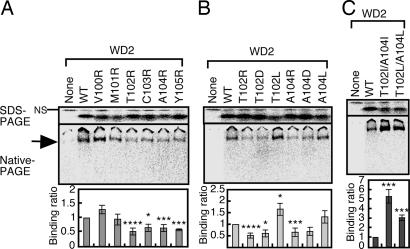
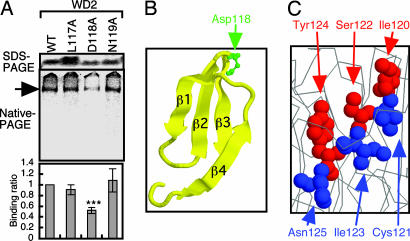
Cytosolic chaperonin containing t-complex polypeptide 1 (CCT)/TRiC is a group II chaperonin that assists in the folding of newly synthesized proteins. It is a eukaryotic homologue of the bacterial group I chaperonin GroEL. In contrast to the well studied functions of GroEL, the substrate recognition mechanism of CCT/TRiC is poorly understood. Here, we established a system for analyzing CCT/TRiC functions by using a reconstituted protein synthesis by using recombinant elements system and show that CCT/TRiC strongly recognizes WD40 proteins particularly at hydrophobic beta-strands. Using the G protein beta subunit (Gbeta), a WD40 protein that is very rich in beta-sheets, as a model substrate, we found that CCT/TRiC prevents aggregation and assists in folding of Gbeta, whereas GroEL does not. Gbeta has a seven-bladed beta-propeller structure; each blade is formed from a WD40 repeat sequence encoding four beta-strands. Detailed mutational analysis of Gbeta indicated that CCT/TRiC, but not GroEL, preferentially recognizes hydrophobic residues aligned on surfaces of beta-strands in the second WD40 repeat of Gbeta. These findings indicate that one of the CCT/TRiC-specific targets is hydrophobic beta-strands, which are highly prone to aggregation.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
Similar articles
-
Domain-specific chaperone-induced expansion is required for beta-actin folding: a comparison of beta-actin conformations upon interactions with GroEL and tail-less complex polypeptide 1 ring complex (TRiC).Biochemistry. 2007 Nov 6;46(44):12639-47. doi: 10.1021/bi700658n. Epub 2007 Oct 16. Biochemistry. 2007. PMID: 17939680
-
Mutational screen identifies critical amino acid residues of beta-actin mediating interaction between its folding intermediates and eukaryotic cytosolic chaperonin CCT.J Struct Biol. 2001 Aug;135(2):185-97. doi: 10.1006/jsbi.2001.4389. J Struct Biol. 2001. PMID: 11580268
-
Newly-synthesized beta-tubulin demonstrates domain-specific interactions with the cytosolic chaperonin.Biochemistry. 1996 Dec 10;35(49):15870-82. doi: 10.1021/bi961114j. Biochemistry. 1996. PMID: 8961952
-
Structure and function of a protein folding machine: the eukaryotic cytosolic chaperonin CCT.FEBS Lett. 2002 Oct 2;529(1):11-6. doi: 10.1016/s0014-5793(02)03180-0. FEBS Lett. 2002. PMID: 12354605 Review.
-
The substrate recognition mechanisms in chaperonins.J Mol Recognit. 2004 Mar-Apr;17(2):85-94. doi: 10.1002/jmr.654. J Mol Recognit. 2004. PMID: 15027029 Review.
Cited by
-
The Mechanism and Function of Group II Chaperonins.J Mol Biol. 2015 Sep 11;427(18):2919-30. doi: 10.1016/j.jmb.2015.04.013. Epub 2015 Apr 30. J Mol Biol. 2015. PMID: 25936650 Free PMC article. Review.
-
MKKS is a centrosome-shuttling protein degraded by disease-causing mutations via CHIP-mediated ubiquitination.Mol Biol Cell. 2008 Mar;19(3):899-911. doi: 10.1091/mbc.e07-07-0631. Epub 2007 Dec 19. Mol Biol Cell. 2008. PMID: 18094050 Free PMC article.
-
The molecular chaperone TRiC/CCT binds to the Trp-Asp 40 (WD40) repeat protein WDR68 and promotes its folding, protein kinase DYRK1A binding, and nuclear accumulation.J Biol Chem. 2014 Nov 28;289(48):33320-32. doi: 10.1074/jbc.M114.586115. Epub 2014 Oct 22. J Biol Chem. 2014. PMID: 25342745 Free PMC article.
-
Defining the TRiC/CCT interactome links chaperonin function to stabilization of newly made proteins with complex topologies.Nat Struct Mol Biol. 2008 Dec;15(12):1255-62. doi: 10.1038/nsmb.1515. Epub 2008 Nov 16. Nat Struct Mol Biol. 2008. PMID: 19011634 Free PMC article.
-
Oligomerization of A. thaliana Heterotrimeric G Protein Subunits AGB1 and AGG2 In Vitro.Protein J. 2020 Oct;39(5):563-573. doi: 10.1007/s10930-020-09914-z. Protein J. 2020. PMID: 32772216
References
-
- Bukau B., Horwich A. L. Cell. 1998;92:351–366. - PubMed
-
- Young J. C., Agashe V. R., Siegers K., Hartl F. U. Nat. Rev. Mol. Cell Biol. 2004;5:781–791. - PubMed
-
- Feltham J. L., Gierasch L. M. Cell. 2000;100:193–196. - PubMed
-
- Saibil H. R., Ranson N. A. Trends Biochem. Sci. 2002;27:627–632. - PubMed
-
- Fenton W. A., Horwich A. L. Q. Rev. Biophys. 2003;36:229–256. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
