

Mba1, a membrane-associated ribosome receptor in mitochondria
- PMID: 16601683
- PMCID: PMC1440829
- DOI: 10.1038/sj.emboj.7601070
Mba1, a membrane-associated ribosome receptor in mitochondria
Abstract
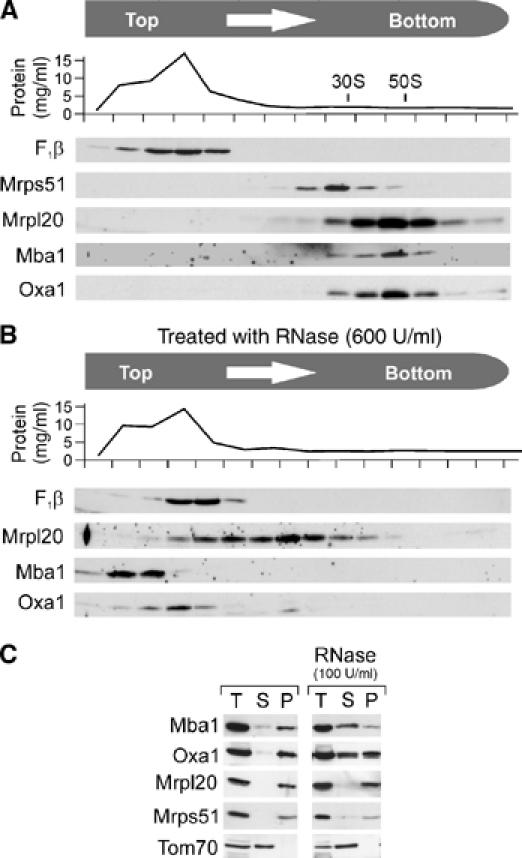
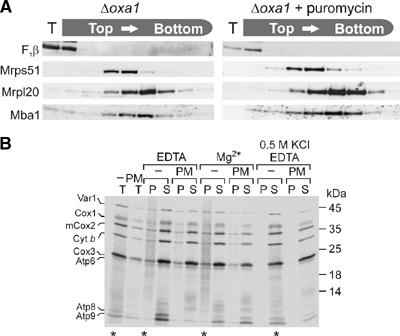
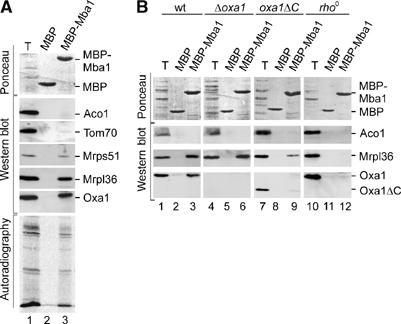
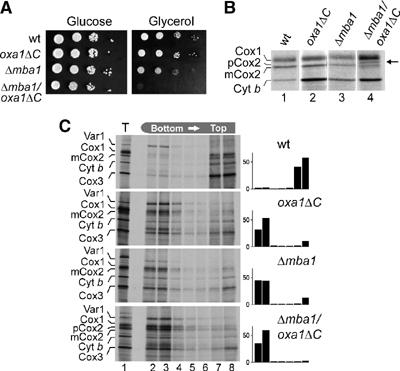
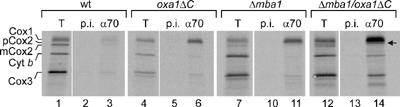
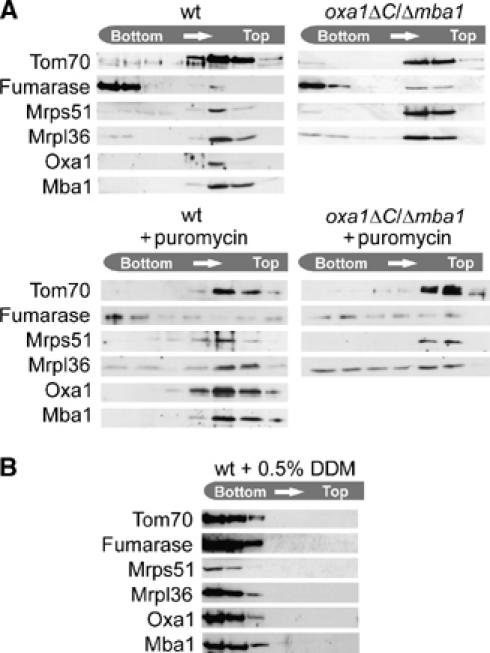
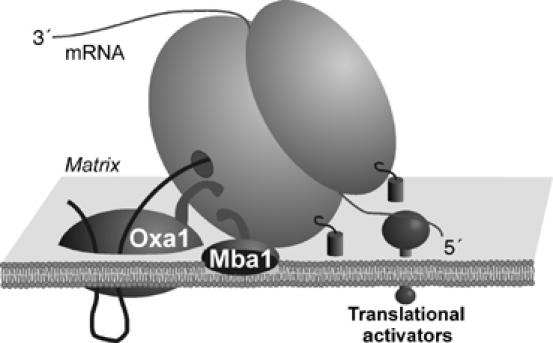
The genome of mitochondria encodes a small number of very hydrophobic polypeptides that are inserted into the inner membrane in a cotranslational reaction. The molecular process by which mitochondrial ribosomes are recruited to the membrane is poorly understood. Here, we show that the inner membrane protein Mba1 binds to the large subunit of mitochondrial ribosomes. It thereby cooperates with the C-terminal ribosome-binding domain of Oxa1, which is a central component of the insertion machinery of the inner membrane. In the absence of both Mba1 and the C-terminus of Oxa1, mitochondrial translation products fail to be properly inserted into the inner membrane and serve as substrates of the matrix chaperone Hsp70. We propose that Mba1 functions as a ribosome receptor that cooperates with Oxa1 in the positioning of the ribosome exit site to the insertion machinery of the inner membrane.
Figures
Similar articles
-
Oxa1-ribosome complexes coordinate the assembly of cytochrome C oxidase in mitochondria.J Biol Chem. 2012 Oct 5;287(41):34484-93. doi: 10.1074/jbc.M112.382630. Epub 2012 Aug 17. J Biol Chem. 2012. PMID: 22904327 Free PMC article.
-
Partial suppression of Oxa1 mutants by mitochondria-targeted signal recognition particle provides insights into the evolution of the cotranslational insertion systems.FEBS J. 2013 Feb;280(3):904-15. doi: 10.1111/febs.12082. Epub 2013 Jan 2. FEBS J. 2013. PMID: 23198851
-
The inner-mitochondrial distribution of Oxa1 depends on the growth conditions and on the availability of substrates.Mol Biol Cell. 2012 Jun;23(12):2292-301. doi: 10.1091/mbc.E11-06-0538. Epub 2012 Apr 18. Mol Biol Cell. 2012. PMID: 22513091 Free PMC article.
-
Co-translational membrane insertion of mitochondrially encoded proteins.Biochim Biophys Acta. 2010 Jun;1803(6):767-75. doi: 10.1016/j.bbamcr.2009.11.010. Epub 2009 Dec 2. Biochim Biophys Acta. 2010. PMID: 19962410 Review.
-
Protein insertion into the inner membrane of mitochondria.IUBMB Life. 2003 Apr-May;55(4-5):219-25. doi: 10.1080/1521654031000123349. IUBMB Life. 2003. PMID: 12880202 Review.
Cited by
-
Oms1 associates with cytochrome c oxidase assembly intermediates to stabilize newly synthesized Cox1.Mol Biol Cell. 2016 May 15;27(10):1570-80. doi: 10.1091/mbc.E15-12-0811. Epub 2016 Mar 30. Mol Biol Cell. 2016. PMID: 27030670 Free PMC article.
-
Oxa1-ribosome complexes coordinate the assembly of cytochrome C oxidase in mitochondria.J Biol Chem. 2012 Oct 5;287(41):34484-93. doi: 10.1074/jbc.M112.382630. Epub 2012 Aug 17. J Biol Chem. 2012. PMID: 22904327 Free PMC article.
-
The process of mammalian mitochondrial protein synthesis.Cell Tissue Res. 2017 Jan;367(1):5-20. doi: 10.1007/s00441-016-2456-0. Epub 2016 Jul 14. Cell Tissue Res. 2017. PMID: 27411691 Free PMC article. Review.
-
Mitochondrial translation and beyond: processes implicated in combined oxidative phosphorylation deficiencies.J Biomed Biotechnol. 2010;2010:737385. doi: 10.1155/2010/737385. Epub 2010 Apr 13. J Biomed Biotechnol. 2010. PMID: 20396601 Free PMC article. Review.
-
Analysis of the assembly profiles for mitochondrial- and nuclear-DNA-encoded subunits into complex I.Mol Cell Biol. 2007 Jun;27(12):4228-37. doi: 10.1128/MCB.00074-07. Epub 2007 Apr 16. Mol Cell Biol. 2007. PMID: 17438127 Free PMC article.
References
-
- Arlt H, Tauer R, Feldmann H, Neupert W, Langer T (1996) The YTA10–12 complex, an AAA protease with chaperone-like activity in the inner membrane of mitochondria. Cell 85: 875–885 - PubMed
-
- Bauer M, Behrens M, Esser K, Michaelis G, Pratje E (1994) PET1402, a nuclear gene required for proteolytic processing of cytochrome oxidase subunit 2 in yeast. Mol Gen Genet 245: 272–278 - PubMed
-
- Beckmann R, Bubeck D, Grassucci R, Penczek P, Verschoor A, Blobel G, Frank J (1997) Alignment of conduits for the nascent polypeptide chain in the ribosome–Sec61 complex. Science 278: 2123–2126 - PubMed
-
- Beckmann R, Spahn CM, Eswar N, Helmers J, Penczek PA, Sali A, Frank J, Blobel G (2001) Architecture of the protein-conducting channel associated with the translating 80S ribosome. Cell 107: 361–372 - PubMed
-
- Bonnefoy N, Chalvet F, Hamel P, Slominski PP, Dujardin G (1994) OXA1, a Saccharomyces cerevisiae nuclear gene whose sequence is conserved from prokaryotes to eukaryotes controls cytochrome oxidase biogenesis. J Mol Biol 239: 201–212 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
