

Coordination of microtubule and microfilament dynamics by Drosophila Rho1, Spire and Cappuccino
- PMID: 16518391
- PMCID: PMC1997291
- DOI: 10.1038/ncb1385
Coordination of microtubule and microfilament dynamics by Drosophila Rho1, Spire and Cappuccino
Abstract
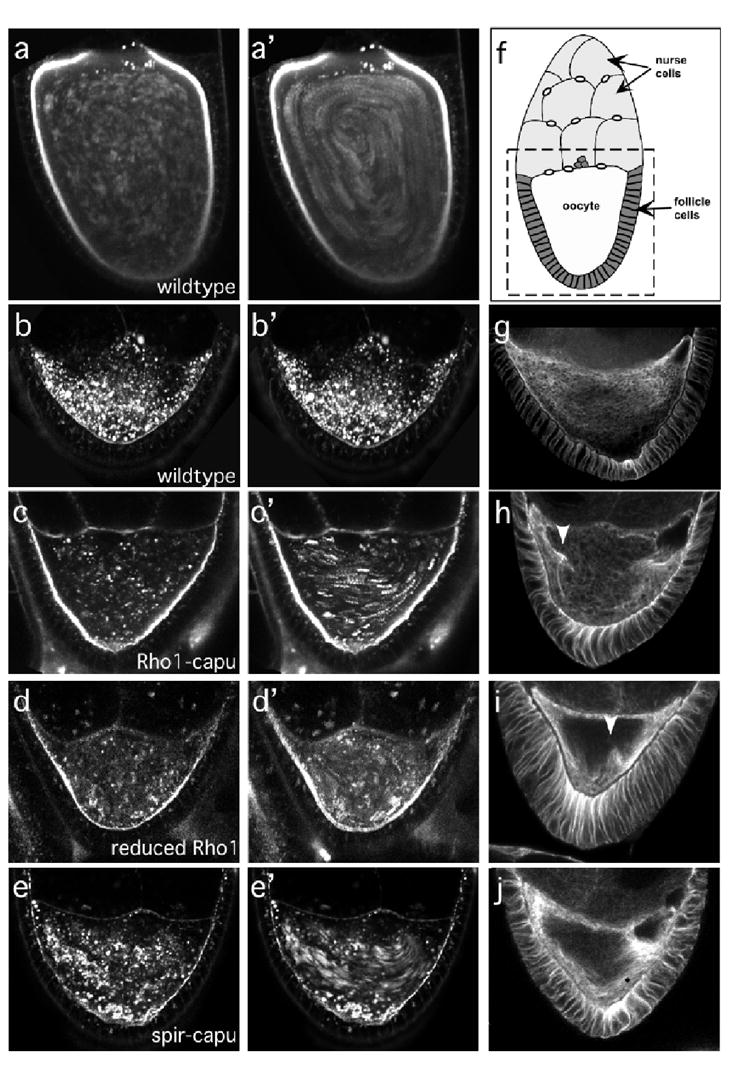
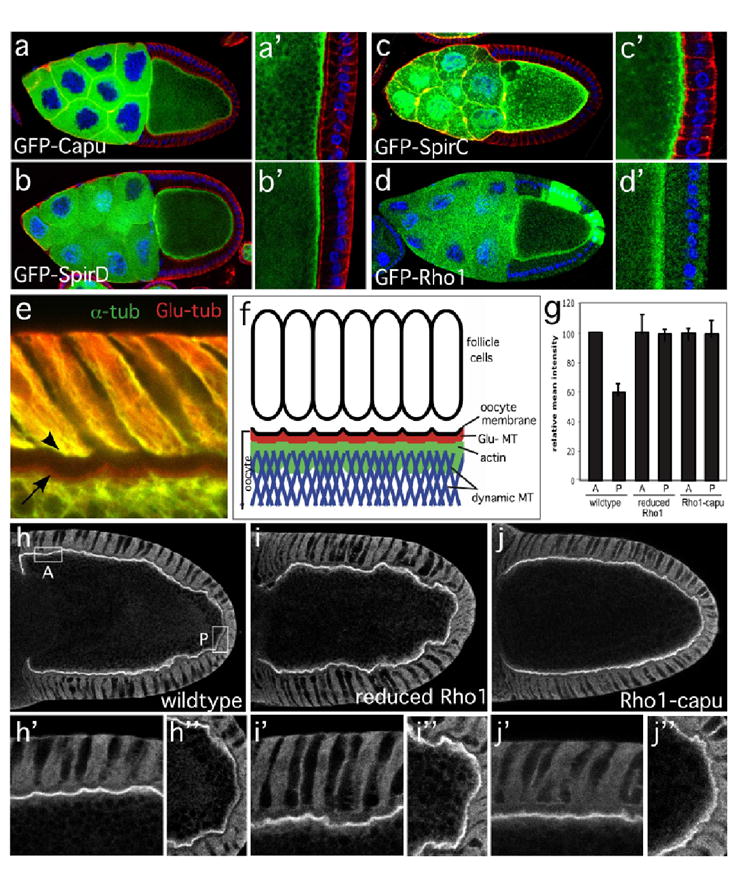
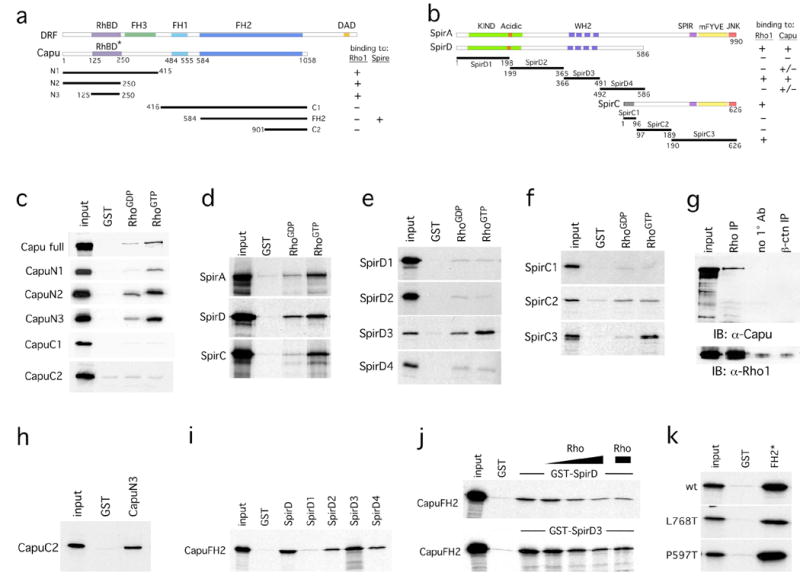
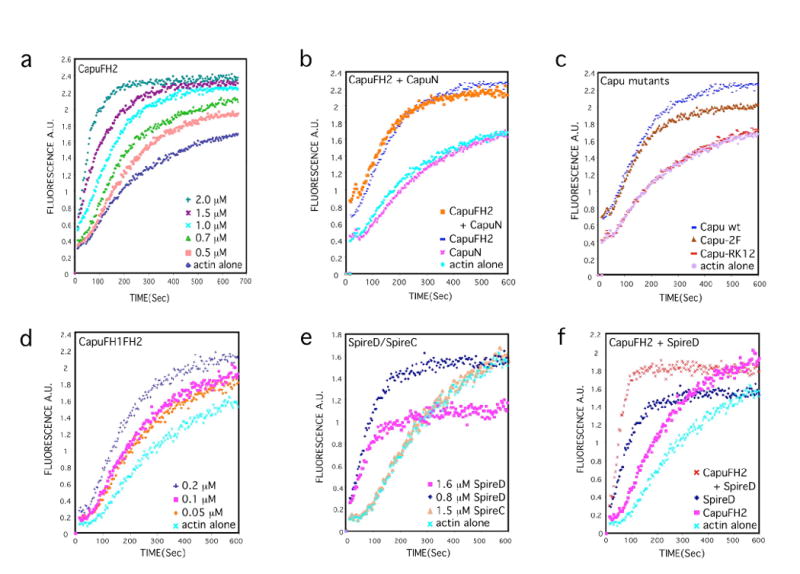
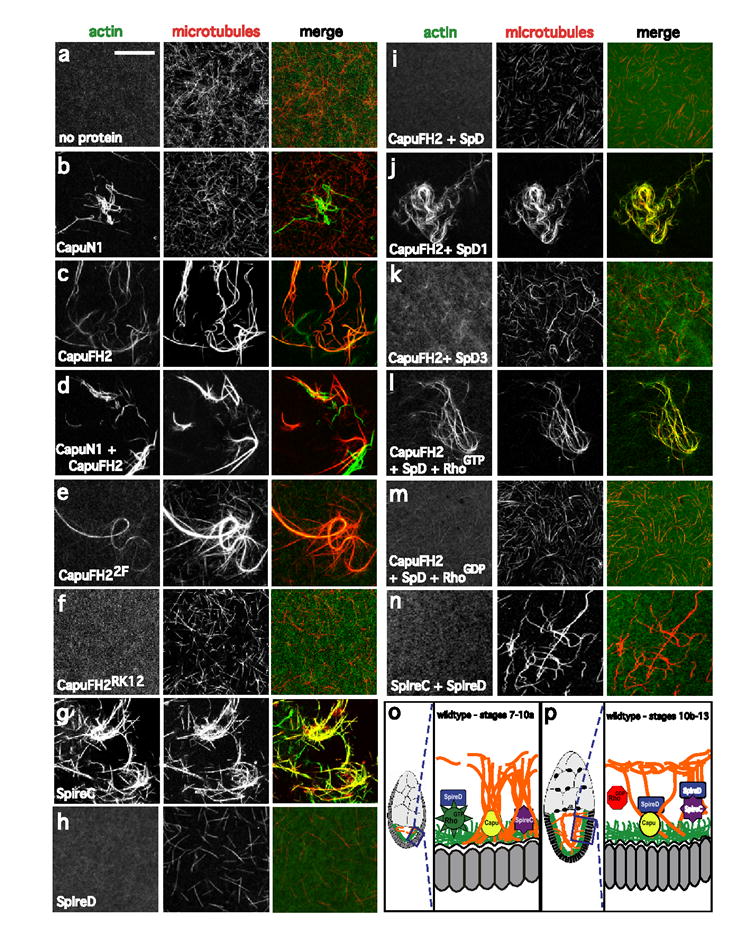
The actin-nucleation factors Spire and Cappuccino (Capu) regulate the onset of ooplasmic streaming in Drosophila melanogaster. Although this streaming event is microtubule-based, actin assembly is required for its timing. It is not understood how the interaction of microtubules and microfilaments is mediated in this context. Here, we demonstrate that Capu and Spire have microtubule and microfilament crosslinking activity. The spire locus encodes several distinct protein isoforms (SpireA, SpireC and SpireD). SpireD was recently shown to nucleate actin, but the activity of the other isoforms has not been addressed. We find that SpireD does not have crosslinking activity, whereas SpireC is a potent crosslinker. We show that SpireD binds to Capu and inhibits F-actin/microtubule crosslinking, and activated Rho1 abolishes this inhibition, establishing a mechanistic basis for the regulation of Capu and Spire activity. We propose that Rho1, cappuccino and spire are elements of a conserved developmental cassette that is capable of directly mediating crosstalk between microtubules and microfilaments.
Figures
Similar articles
-
Interaction between microtubules and the Drosophila formin Cappuccino and its effect on actin assembly.J Biol Chem. 2014 Feb 14;289(7):4395-404. doi: 10.1074/jbc.M113.499921. Epub 2013 Dec 20. J Biol Chem. 2014. PMID: 24362037 Free PMC article.
-
Regulatory interactions between two actin nucleators, Spire and Cappuccino.J Cell Biol. 2007 Oct 8;179(1):117-28. doi: 10.1083/jcb.200706196. J Cell Biol. 2007. PMID: 17923532 Free PMC article.
-
Capu and Spire assemble a cytoplasmic actin mesh that maintains microtubule organization in the Drosophila oocyte.Dev Cell. 2007 Oct;13(4):539-53. doi: 10.1016/j.devcel.2007.09.003. Dev Cell. 2007. PMID: 17925229 Free PMC article.
-
Cellular functions of the Spir actin-nucleation factors.Trends Cell Biol. 2006 Sep;16(9):477-83. doi: 10.1016/j.tcb.2006.07.005. Epub 2006 Aug 9. Trends Cell Biol. 2006. PMID: 16901698 Review.
-
[Advances in the study of the actin nucleation factor Spire].Sheng Li Xue Bao. 2024 Apr 25;76(2):341-345. Sheng Li Xue Bao. 2024. PMID: 38658382 Review. Chinese.
Cited by
-
Formins in development: orchestrating body plan origami.Biochim Biophys Acta. 2010 Feb;1803(2):207-25. doi: 10.1016/j.bbamcr.2008.09.016. Epub 2008 Oct 14. Biochim Biophys Acta. 2010. PMID: 18996154 Free PMC article. Review.
-
Cytoplasmic diffusion: molecular motors mix it up.J Cell Biol. 2008 Nov 17;183(4):583-7. doi: 10.1083/jcb.200806149. Epub 2008 Nov 10. J Cell Biol. 2008. PMID: 19001127 Free PMC article. Review.
-
Coordinating Proliferation, Polarity, and Cell Fate in the Drosophila Female Germline.Front Cell Dev Biol. 2020 Feb 4;8:19. doi: 10.3389/fcell.2020.00019. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32117961 Free PMC article. Review.
-
Formins and microtubules.Biochim Biophys Acta. 2010 Feb;1803(2):164-73. doi: 10.1016/j.bbamcr.2009.07.006. Epub 2009 Jul 23. Biochim Biophys Acta. 2010. PMID: 19631698 Free PMC article. Review.
-
Structures of actin-bound Wiskott-Aldrich syndrome protein homology 2 (WH2) domains of Spire and the implication for filament nucleation.Proc Natl Acad Sci U S A. 2010 Jun 29;107(26):11757-62. doi: 10.1073/pnas.1005347107. Epub 2010 Jun 10. Proc Natl Acad Sci U S A. 2010. PMID: 20538977 Free PMC article.
References
-
- Theurkauf WE. Premature microtubule-dependent cytoplasmic streaming in cappuccino and spire mutant oocytes. Science. 1994;265:2093–2096. - PubMed
-
- Manseau L, Schüpbach T. cappuccino and spire: two unique maternal-effect loci required for both the anteroposterior and dorsoventral patterns of the Drosophila embryo. Genes Dev. 1989;3:1437–1452. - PubMed
-
- Emmons S, et al. cappuccino, a Drosophila maternal effect gene required for polarity of the egg and embryo, is related to the vertebrate limb deformity locus. Genes Dev. 1995;9:2482–2494. - PubMed
-
- Wellington A, et al. Spire contains actin binding domains and is related to ascidian posterior end mark-5. Development. 1999;126:5267–74. - PubMed
-
- Cooley L, Theurkauf WE. Cytoskeletal functions during Drosophila oogenesis. Science. 1994;266:590–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
