

Molecular and morphological heterogeneity of neural precursors in the mouse neocortical proliferative zones
- PMID: 16421324
- PMCID: PMC3249619
- DOI: 10.1523/JNEUROSCI.4499-05.2006
Molecular and morphological heterogeneity of neural precursors in the mouse neocortical proliferative zones
Abstract
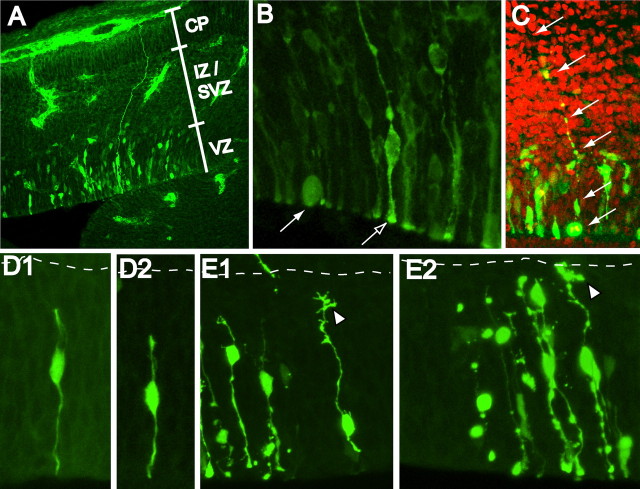
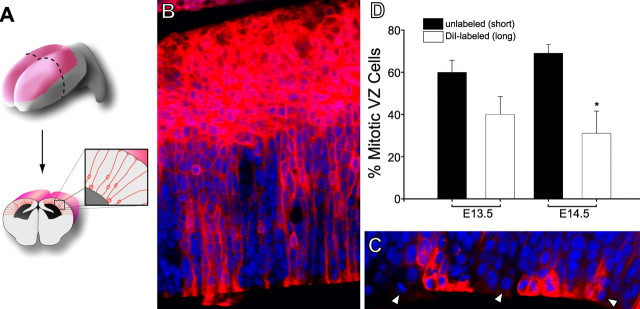
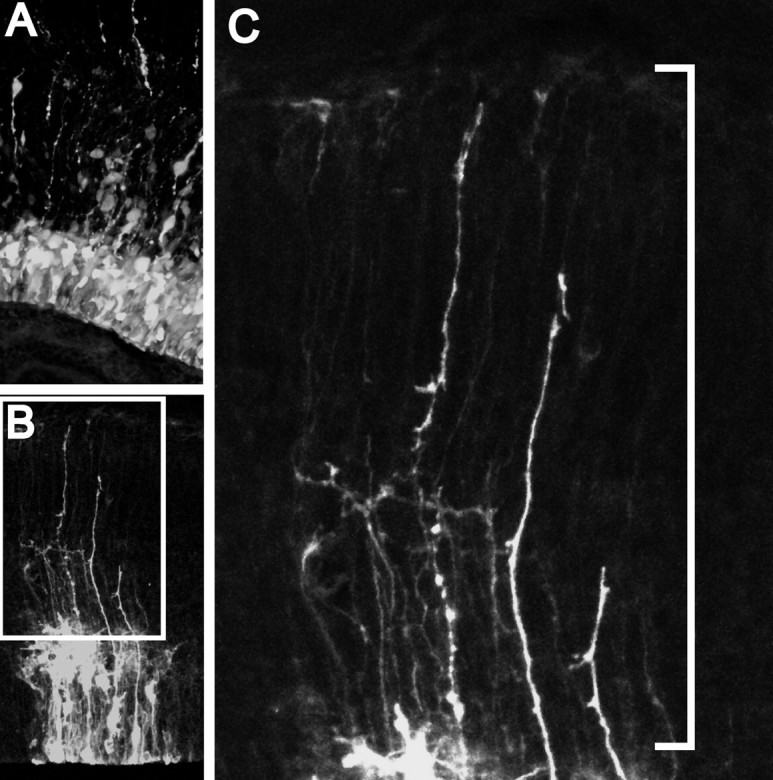
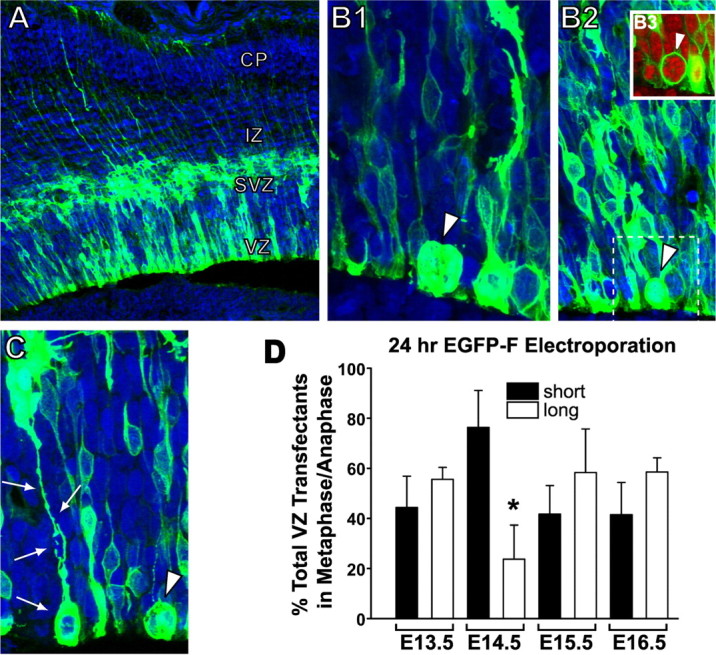
The proliferative ventricular zone (VZ) is the main source of projection neurons for the overlying cerebral neocortex. The number and diversity of neocortical neurons is determined, in part, by factors controlling the proliferation and specification of VZ cells during embryonic development. We used a variety of methods, including in utero electroporation with specific cellular markers, computer-assisted serial EM cell reconstruction, and time-lapse multiphoton imaging to characterize the molecular and morphological characteristics of the VZ constituents and to capture their behavior during cell division. Our analyses reveal at least two types of dividing cells in the VZ: (1) radial glial cells (RGCs) that span the entire neocortical wall and maintain contact both at the ventricular and pial surfaces throughout mitotic division, and (2) short neural precursors (SNPs) that possess a ventricular endfoot and a basal process of variable length that is retracted during mitotic division. These two precursor cell classes are present concomitantly in the VZ, but their relative number changes over the course of cortical neurogenesis. Moreover, the SNPs are morphologically, ultrastructurally and molecularly distinct from dividing RGCs. For example, SNPs are marked by their preferential expression of the tubulin alpha-1 promoter whereas RGCs instead express the glutamate-aspartate transporter and brain lipid binding protein promoters. In contrast to recent studies that suggest that RGCs are the sole type of VZ precursor, the present study indicates that the VZ in murine dorsal telencephalon is similar to that in human and nonhuman primates, because it contains multiple types of neuronal precursors.
Figures
Similar articles
-
Heterogeneity in ventricular zone neural precursors contributes to neuronal fate diversity in the postnatal neocortex.J Neurosci. 2010 May 19;30(20):7028-36. doi: 10.1523/JNEUROSCI.6131-09.2010. J Neurosci. 2010. PMID: 20484645 Free PMC article.
-
beta1 integrin maintains integrity of the embryonic neocortical stem cell niche.PLoS Biol. 2009 Aug;7(8):e1000176. doi: 10.1371/journal.pbio.1000176. Epub 2009 Aug 18. PLoS Biol. 2009. PMID: 19688041 Free PMC article.
-
Diversity of neural precursor cell types in the prenatal macaque cerebral cortex exists largely within the astroglial cell lineage.PLoS One. 2013 May 28;8(5):e63848. doi: 10.1371/journal.pone.0063848. Print 2013. PLoS One. 2013. PMID: 23724007 Free PMC article.
-
Calcium signaling in neocortical development.Dev Neurobiol. 2015 Apr;75(4):360-8. doi: 10.1002/dneu.22273. Epub 2015 Feb 18. Dev Neurobiol. 2015. PMID: 25652687 Review.
-
Cytoarchitecture of mouse and human subventricular zone in developing cerebral neocortex.Exp Brain Res. 2012 Jan;216(2):161-8. doi: 10.1007/s00221-011-2933-3. Epub 2011 Nov 13. Exp Brain Res. 2012. PMID: 22080150 Free PMC article. Review.
Cited by
-
GRM7 regulates embryonic neurogenesis via CREB and YAP.Stem Cell Reports. 2015 May 12;4(5):795-810. doi: 10.1016/j.stemcr.2015.03.004. Epub 2015 Apr 23. Stem Cell Reports. 2015. PMID: 25921811 Free PMC article.
-
Shaping our minds: stem and progenitor cell diversity in the mammalian neocortex.Neuron. 2013 Jan 9;77(1):19-34. doi: 10.1016/j.neuron.2012.12.022. Neuron. 2013. PMID: 23312513 Free PMC article. Review.
-
Coevolution of radial glial cells and the cerebral cortex.Glia. 2015 Aug;63(8):1303-19. doi: 10.1002/glia.22827. Epub 2015 Mar 23. Glia. 2015. PMID: 25808466 Free PMC article. Review.
-
The Long and the Short of it: Gene and Environment Interactions During Early Cortical Development and Consequences for Long-Term Neurological Disease.Front Psychiatry. 2012 Jun 12;3:50. doi: 10.3389/fpsyt.2012.00050. eCollection 2012. Front Psychiatry. 2012. PMID: 22701439 Free PMC article.
-
Tbr2-positive intermediate (basal) neuronal progenitors safeguard cerebral cortex expansion by controlling amplification of pallial glutamatergic neurons and attraction of subpallial GABAergic interneurons.Genes Dev. 2010 Aug 15;24(16):1816-26. doi: 10.1101/gad.575410. Genes Dev. 2010. PMID: 20713522 Free PMC article.
References
-
- Akamatsu W, Okano HJ, Osumi N, Inoue T, Nakamura S, Sakakibara S, Miura M, Matsuo N, Darnell RB, Okano H (1999) Mammalian ELAV-like neuronal RNA-binding proteins HuB and HuC promote neuronal development in both the central and the peripheral nervous systems. Proc Natl Acad Sci USA 96: 9885–9890. - PMC - PubMed
-
- Angevine JB, Sidman RL (1961) Autoradiographic study of cell migration during histogenesis of the cerebral cortex in the mouse. Nature 192: 766–768. - PubMed
-
- Anthony TE, Klein C, Fishell G, Heintz N (2004) Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron 41: 881–890. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical