

PTF1 is an organ-specific and Notch-independent basic helix-loop-helix complex containing the mammalian Suppressor of Hairless (RBP-J) or its paralogue, RBP-L
- PMID: 16354684
- PMCID: PMC1317634
- DOI: 10.1128/MCB.26.1.117-130.2006
PTF1 is an organ-specific and Notch-independent basic helix-loop-helix complex containing the mammalian Suppressor of Hairless (RBP-J) or its paralogue, RBP-L
Abstract
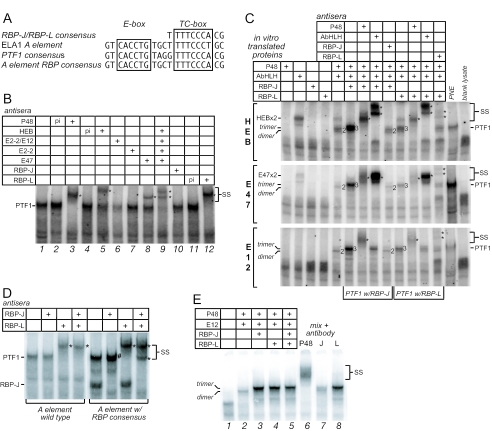
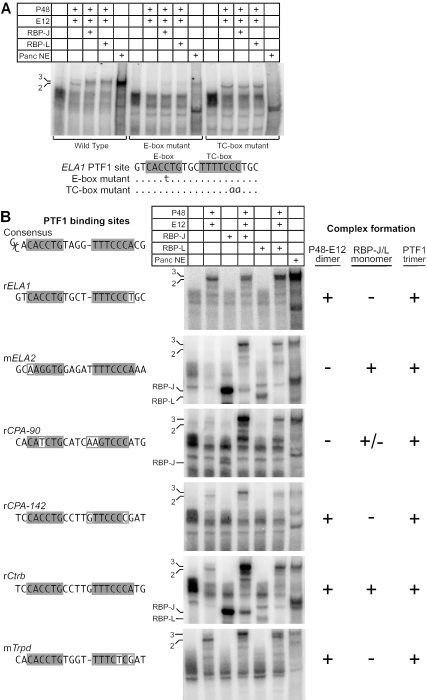
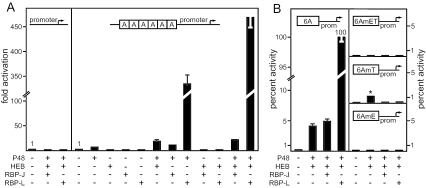
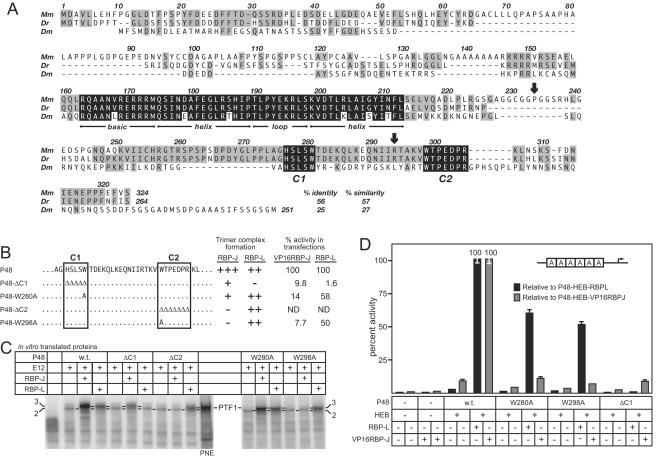
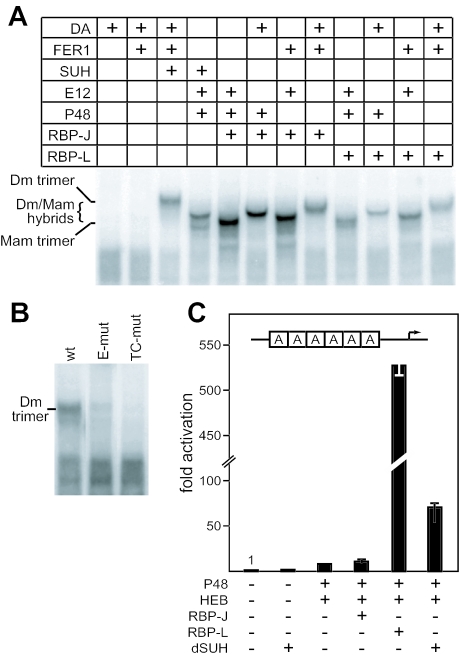
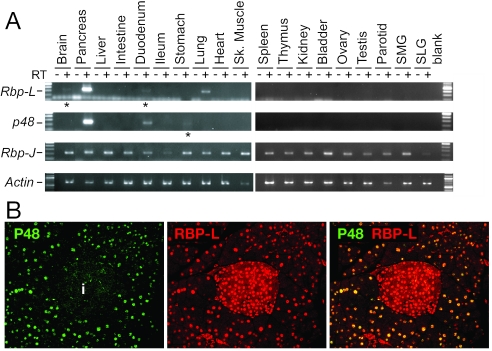
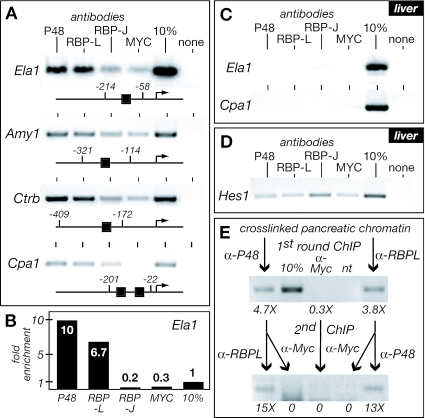
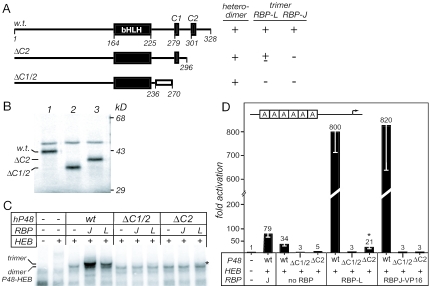
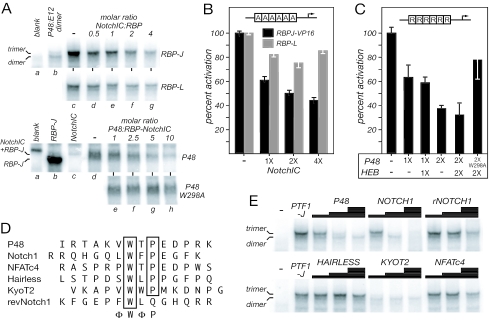
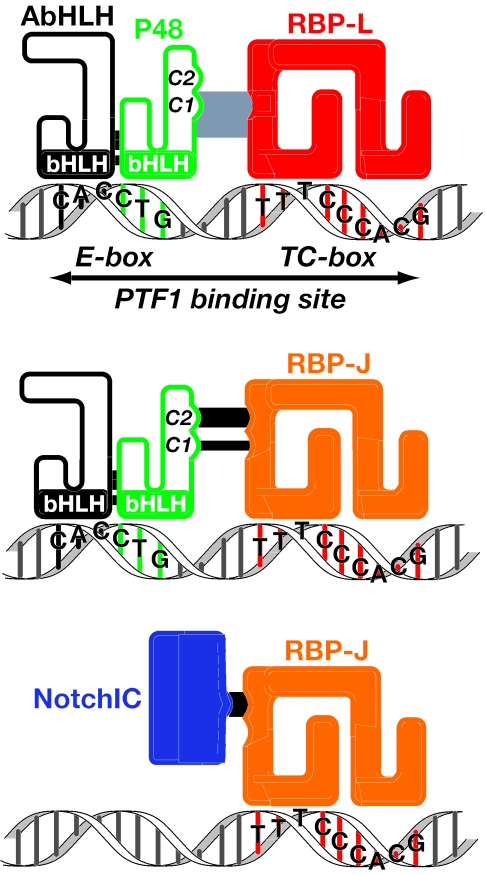
PTF1 is a trimeric transcription factor essential to the development of the pancreas and to the maintenance of the differentiated state of the adult exocrine pancreas. It comprises a dimer of P48/PTF1a (a pancreas and neural restricted basic helix-loop-helix [bHLH] protein) and a class A bHLH protein, together with a third protein that we show can be either the mammalian Suppressor of Hairless (RBP-J) or its paralogue, RBP-L. In mature acinar cells, PTF1 exclusively contains the RBP-L isoform and is bound to the promoters of acinar specific genes. P48 interacts with the RBP subunit primarily through two short conserved tryptophan-containing motifs, similar to the motif of the Notch intracellular domain (NotchIC) that interacts with RBP-J. The transcriptional activities of the J and L forms of PTF1 are independent of Notch signaling, because P48 occupies the NotchIC docking site on RBP-J and RBP-L does not bind the NotchIC. Mutations that delete one or both of the RBP-interacting motifs of P48 eliminate RBP-binding and are associated with a human genetic disorder characterized by pancreatic and cerebellar agenesis, which indicates that the association of P48 and RBPs is required for proper embryonic development. The presence of related peptide motifs in other transcription factors indicates a broader Notch-independent function for RBPJ/SU(H).
Figures
Similar articles
-
Early pancreatic development requires the vertebrate Suppressor of Hairless (RBPJ) in the PTF1 bHLH complex.Genes Dev. 2007 Oct 15;21(20):2629-43. doi: 10.1101/gad.1575207. Genes Dev. 2007. PMID: 17938243 Free PMC article.
-
p48 subunit of mouse PTF1 binds to RBP-Jkappa/CBF-1, the intracellular mediator of Notch signalling, and is expressed in the neural tube of early stage embryos.Genes Cells. 2001 Apr;6(4):345-60. doi: 10.1046/j.1365-2443.2001.00422.x. Genes Cells. 2001. PMID: 11318877
-
p/CAF modulates the activity of the transcription factor p48/Ptf1a involved in pancreatic acinar differentiation.Biochem J. 2009 Mar 1;418(2):463-73. doi: 10.1042/BJ20080293. Biochem J. 2009. PMID: 18834332
-
Two opposing roles of RBP-J in Notch signaling.Curr Top Dev Biol. 2010;92:231-52. doi: 10.1016/S0070-2153(10)92007-3. Curr Top Dev Biol. 2010. PMID: 20816397 Review.
-
The effects of conformational heterogeneity on the binding of the Notch intracellular domain to effector proteins: a case of biologically tuned disorder.Biochem Soc Trans. 2008 Apr;36(Pt 2):157-66. doi: 10.1042/BST0360157. Biochem Soc Trans. 2008. PMID: 18363556 Review.
Cited by
-
TNF-induced osteoclastogenesis and inflammatory bone resorption are inhibited by transcription factor RBP-J.J Exp Med. 2012 Feb 13;209(2):319-34. doi: 10.1084/jem.20111566. Epub 2012 Jan 16. J Exp Med. 2012. PMID: 22249448 Free PMC article.
-
Differential ability of Ptf1a and Ptf1a-VP16 to convert stomach, duodenum and liver to pancreas.Dev Biol. 2007 Apr 15;304(2):786-99. doi: 10.1016/j.ydbio.2007.01.027. Epub 2007 Jan 25. Dev Biol. 2007. PMID: 17320068 Free PMC article.
-
Functional and association analysis of an Amerindian-derived population-specific p.(Thr280Met) variant in RBPJL, a component of the PTF1 complex.Eur J Hum Genet. 2018 Feb;26(2):238-246. doi: 10.1038/s41431-017-0062-6. Epub 2018 Jan 4. Eur J Hum Genet. 2018. PMID: 29302047 Free PMC article.
-
Neonatal diabetes mellitus.Endocr Rev. 2008 May;29(3):265-91. doi: 10.1210/er.2007-0029. Epub 2008 Apr 24. Endocr Rev. 2008. PMID: 18436707 Free PMC article. Review.
-
Transcriptional control of mammalian pancreas organogenesis.Cell Mol Life Sci. 2014 Jul;71(13):2383-402. doi: 10.1007/s00018-013-1510-2. Epub 2013 Nov 13. Cell Mol Life Sci. 2014. PMID: 24221136 Free PMC article. Review.
References
-
- Apelqvist, A., H. Li, S. L., P. Beatus, D. J. Anderson, T. Honjo, M. Hrabe de Angelis, U. Lendahl, and H. Edlund. 1999. Notch signalling controls pancreatic cell differentiation. Nature 400:877-881. - PubMed
-
- Artavanis-Tsakonas, S., M. D. Rand, and R. J. Lake. 1999. Notch signaling: cell fate control and signal integration in development. Science 284:770-776. - PubMed
-
- Barolo, S., R. G. Walker, A. D. Polyanovsky, G. Freschi, T. Keil, and J. W. Posakony. 2000. A Notch-independent activity of Suppressor of Hairless is required for normal mechanoreceptor physiology. Cell 103:957-969. - PubMed
-
- Blackwell, T. K., and H. Weintraub. 1990. Differences and similarities in DNA-binding preferences of MyoD and E2A protein complexes revealed by binding site selection. Science 250:1104-1110. - PubMed
-
- Chaya, D., and K. S. Zaret. 2003. Sequential chromatin immunoprecipitation from animal tissues. Methods Enzymol. 376:361-372. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases