

ABA-hypersensitive germination3 encodes a protein phosphatase 2C (AtPP2CA) that strongly regulates abscisic acid signaling during germination among Arabidopsis protein phosphatase 2Cs
- PMID: 16339800
- PMCID: PMC1326036
- DOI: 10.1104/pp.105.070128
ABA-hypersensitive germination3 encodes a protein phosphatase 2C (AtPP2CA) that strongly regulates abscisic acid signaling during germination among Arabidopsis protein phosphatase 2Cs
Abstract
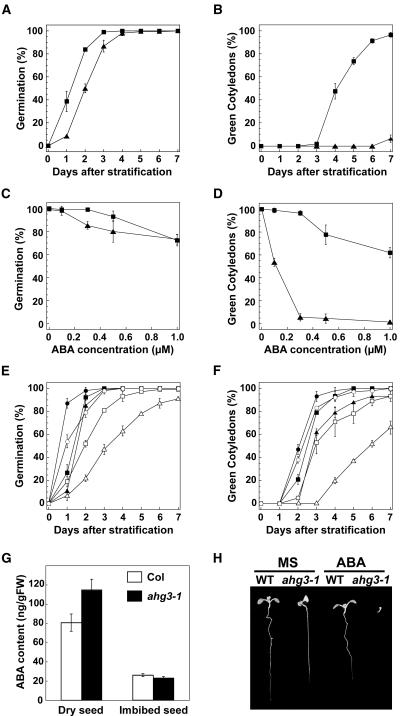
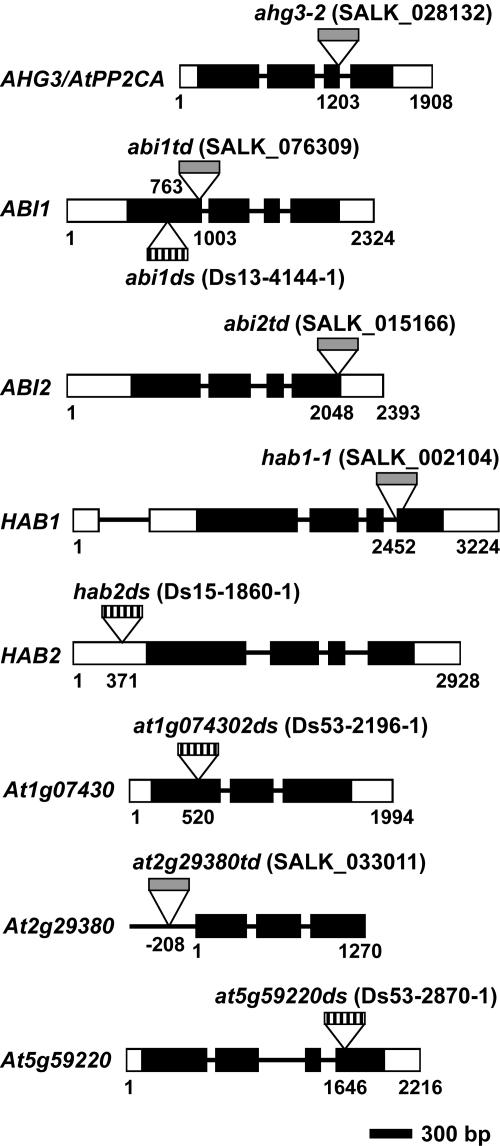
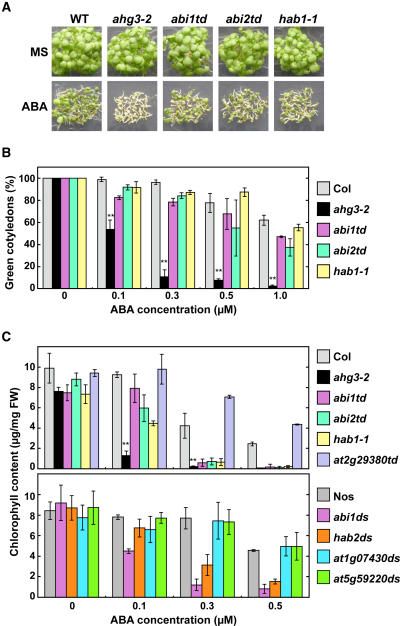
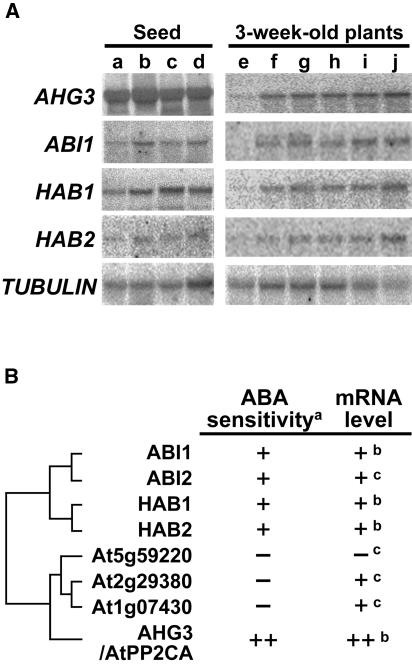
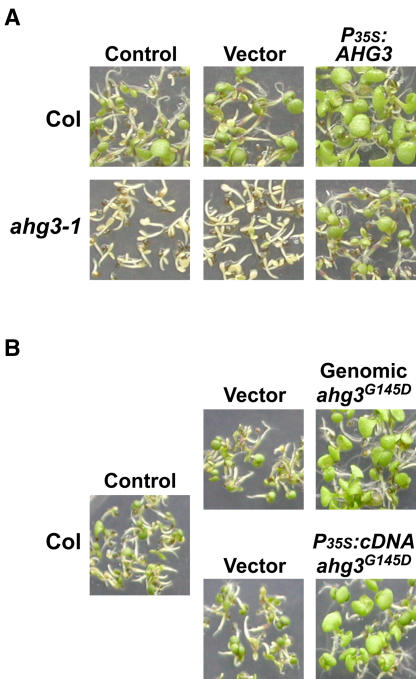
The phytohormone abscisic acid (ABA) regulates physiologically important developmental processes and stress responses. Previously, we reported on Arabidopsis (Arabidopsis thaliana) L. Heynh. ahg mutants, which are hypersensitive to ABA during germination and early growth. Among them, ABA-hypersensitive germination3 (ahg3) showed the strongest ABA hypersensitivity. In this study, we found that the AHG3 gene is identical to AtPP2CA, which encodes a protein phosphatase 2C (PP2C). Although AtPP2CA has been reported to be involved in the ABA response on the basis of results obtained by reverse-genetics approaches, its physiological relevance in the ABA response has not been clarified yet. We demonstrate in vitro and in vivo that the ahg3-1 missense mutation causes the loss of PP2C activity, providing concrete confirmation that this PP2C functions as a negative regulator in ABA signaling. Furthermore, we compared the effects of disruption mutations of eight structurally related PP2C genes of Arabidopsis, including ABI1, ABI2, HAB1, and HAB2, and found that the disruptant mutant of AHG3/AtPP2CA had the strongest ABA hypersensitivity during germination, but it did not display any significant phenotypes in adult plants. Northern-blot analysis clearly showed that AHG3/AtPP2CA is the most active among those PP2C genes in seeds. These results suggest that AHG3/AtPP2CA plays a major role among PP2Cs in the ABA response in seeds and that the functions of those PP2Cs overlap, but their unique tissue- or development-specific expression confers distinct and indispensable physiological functions in the ABA response.
Figures

Similar articles
-
The protein phosphatase AtPP2CA negatively regulates abscisic acid signal transduction in Arabidopsis, and effects of abh1 on AtPP2CA mRNA.Plant Physiol. 2006 Jan;140(1):127-39. doi: 10.1104/pp.105.070318. Epub 2005 Dec 16. Plant Physiol. 2006. PMID: 16361522 Free PMC article.
-
ABA-Hypersensitive Germination1 encodes a protein phosphatase 2C, an essential component of abscisic acid signaling in Arabidopsis seed.Plant J. 2007 Jun;50(6):935-49. doi: 10.1111/j.1365-313X.2007.03107.x. Epub 2007 Apr 25. Plant J. 2007. PMID: 17461784
-
Modulation of abscisic acid signaling in vivo by an engineered receptor-insensitive protein phosphatase type 2C allele.Plant Physiol. 2011 May;156(1):106-16. doi: 10.1104/pp.110.170894. Epub 2011 Feb 28. Plant Physiol. 2011. PMID: 21357183 Free PMC article.
-
Protein phosphatase 2C (PP2C) function in higher plants.Plant Mol Biol. 1998 Dec;38(6):919-27. doi: 10.1023/a:1006054607850. Plant Mol Biol. 1998. PMID: 9869399 Review.
-
Regulation of Arabidopsis MAPKKK18 by ABI1 and SnRK2, components of the ABA signaling pathway.Plant Signal Behav. 2016;11(4):e1139277. doi: 10.1080/15592324.2016.1139277. Plant Signal Behav. 2016. PMID: 26852793 Free PMC article. Review.
Cited by
-
Genome-wide characterization and expression analysis of TOPP-type protein phosphatases in soybean (Glycine max L.) reveal the role of GmTOPP13 in drought tolerance.Genes Genomics. 2021 Jul;43(7):783-796. doi: 10.1007/s13258-021-01075-2. Epub 2021 Apr 17. Genes Genomics. 2021. PMID: 33864615
-
Sequence Polymorphisms at the REDUCED DORMANCY5 Pseudophosphatase Underlie Natural Variation in Arabidopsis Dormancy.Plant Physiol. 2016 Aug;171(4):2659-70. doi: 10.1104/pp.16.00525. Epub 2016 Jun 10. Plant Physiol. 2016. PMID: 27288362 Free PMC article.
-
Selective inhibition of clade A phosphatases type 2C by PYR/PYL/RCAR abscisic acid receptors.Plant Physiol. 2012 Feb;158(2):970-80. doi: 10.1104/pp.111.188623. Epub 2011 Dec 23. Plant Physiol. 2012. PMID: 22198272 Free PMC article.
-
Overexpression of a protein phosphatase 2C from beech seeds in Arabidopsis shows phenotypes related to abscisic acid responses and gibberellin biosynthesis.Plant Physiol. 2006 Aug;141(4):1414-24. doi: 10.1104/pp.106.084681. Epub 2006 Jun 30. Plant Physiol. 2006. PMID: 16815952 Free PMC article.
-
Identification and mechanism of ABA receptor antagonism.Nat Struct Mol Biol. 2010 Sep;17(9):1102-8. doi: 10.1038/nsmb.1887. Epub 2010 Aug 22. Nat Struct Mol Biol. 2010. PMID: 20729862 Free PMC article.
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796–815 - PubMed
-
- Asami T, Sekimata K, Wang JM, Yoneyama K, Takeuchi Y, Yoshida S (1999) Preparation of (±)-[1,2-13C2]abscisic acid for use as a stable and pure internal standard. J Chem Res Synop 11: 658–659
-
- Bertauche N, Leung J, Giraudat J (1996) Protein phosphatase activity of abscisic acid insensitive 1 (ABI1) protein from Arabidopsis thaliana. Eur J Biochem 241: 193–200 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
