

CD36 involvement in orosensory detection of dietary lipids, spontaneous fat preference, and digestive secretions
- PMID: 16276419
- PMCID: PMC1265871
- DOI: 10.1172/JCI25299
CD36 involvement in orosensory detection of dietary lipids, spontaneous fat preference, and digestive secretions
Abstract
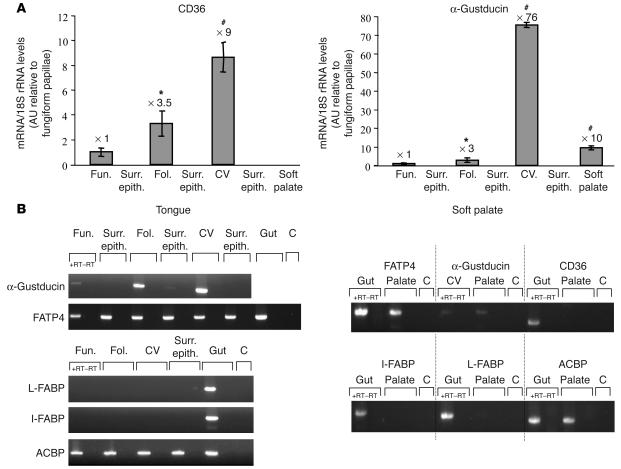
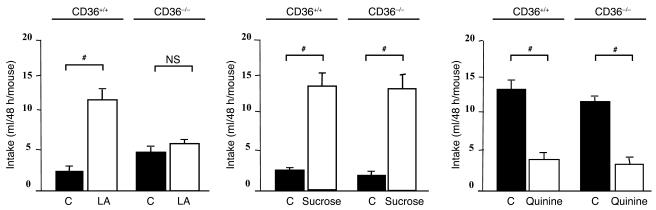
Rats and mice exhibit a spontaneous attraction for lipids. Such a behavior raises the possibility that an orosensory system is responsible for the detection of dietary lipids. The fatty acid transporter CD36 appears to be a plausible candidate for this function since it has a high affinity for long-chain fatty acids (LCFAs) and is found in lingual papillae in the rat. To explore this hypothesis further, experiments were conducted in rats and in wild-type and CD36-null mice. In mice, RT-PCR experiments with primers specific for candidate lipid-binding proteins revealed that only CD36 expression was restricted to lingual papillae although absent from the palatal papillae. Immunostaining studies showed a distribution of CD36 along the apical side of circumvallate taste bud cells. CD36 gene inactivation fully abolished the preference for LCFA-enriched solutions and solid diet observed in wild-type mice. Furthermore, in rats and wild-type mice with an esophageal ligation, deposition of unsaturated LCFAs onto the tongue led to a rapid and sustained rise in flux and protein content of pancreatobiliary secretions. These findings demonstrate that CD36 is involved in oral LCFA detection and raise the possibility that an alteration in the lingual fat perception may be linked to feeding dysregulation.
Figures





Comment in
-
CD36 may determine our desire for dietary fats.J Clin Invest. 2005 Nov;115(11):2965-7. doi: 10.1172/JCI26955. J Clin Invest. 2005. PMID: 16276408 Free PMC article. Review.
Similar articles
-
Apical CD36 immunolocalization in human and porcine taste buds from circumvallate and foliate papillae.Acta Histochem. 2011 Dec;113(8):839-43. doi: 10.1016/j.acthis.2010.08.006. Epub 2010 Oct 14. Acta Histochem. 2011. PMID: 20950842
-
Cell mechanisms of gustatory lipids perception and modulation of the dietary fat preference.Biochimie. 2014 Dec;107 Pt A:11-4. doi: 10.1016/j.biochi.2014.06.018. Epub 2014 Jul 2. Biochimie. 2014. PMID: 24997404 Review.
-
The lipid-sensor candidates CD36 and GPR120 are differentially regulated by dietary lipids in mouse taste buds: impact on spontaneous fat preference.PLoS One. 2011;6(8):e24014. doi: 10.1371/journal.pone.0024014. Epub 2011 Aug 25. PLoS One. 2011. PMID: 21901153 Free PMC article.
-
The gustatory pathway is involved in CD36-mediated orosensory perception of long-chain fatty acids in the mouse.FASEB J. 2008 May;22(5):1458-68. doi: 10.1096/fj.07-8415com. Epub 2007 Dec 27. FASEB J. 2008. PMID: 18162488
-
CD36 and taste of fat.Curr Opin Clin Nutr Metab Care. 2012 Mar;15(2):107-11. doi: 10.1097/MCO.0b013e32834ff19c. Curr Opin Clin Nutr Metab Care. 2012. PMID: 22248592 Review.
Cited by
-
The Macronutrients, Appetite, and Energy Intake.Annu Rev Nutr. 2016 Jul 17;36:73-103. doi: 10.1146/annurev-nutr-121415-112624. Annu Rev Nutr. 2016. PMID: 27431364 Free PMC article. Review.
-
Saliva and other taste stimuli are important for gustatory processing of linoleic acid.Am J Physiol Regul Integr Comp Physiol. 2009 Oct;297(4):R1162-70. doi: 10.1152/ajpregu.00217.2009. Epub 2009 Aug 19. Am J Physiol Regul Integr Comp Physiol. 2009. PMID: 19692663 Free PMC article.
-
Taste preference for fatty acids is mediated by GPR40 and GPR120.J Neurosci. 2010 Jun 23;30(25):8376-82. doi: 10.1523/JNEUROSCI.0496-10.2010. J Neurosci. 2010. PMID: 20573884 Free PMC article.
-
CD36 - A novel molecular target in the neurovascular unit.Eur J Neurosci. 2021 Apr;53(8):2500-2510. doi: 10.1111/ejn.15147. Epub 2021 Feb 28. Eur J Neurosci. 2021. PMID: 33560561 Free PMC article. Review.
-
From appetite setpoint to appetition: 50years of ingestive behavior research.Physiol Behav. 2018 Aug 1;192:210-217. doi: 10.1016/j.physbeh.2018.01.001. Epub 2018 Jan 2. Physiol Behav. 2018. PMID: 29305256 Free PMC article. Review.
References
-
- Drewnowski A, Brunzell JD, Sande K, Iverius PH, Greenwood MR. Sweet tooth reconsidered: taste responsiveness in human obesity. Physiol. Behav. 1985;35:617–622. - PubMed
-
- Mela DJ, Sacchetti DA. Sensory preferences for fats: relationships with diet and body composition. Am. J. Clin. Nutr. 1991;53:908–915. - PubMed
-
- French S, Robinson T. Fats and food intake. Curr. Opin. Clin. Nutr. Metab. Care. 2003;6:629–634. - PubMed
-
- Mela DJ. Sensory assessment of fat content in fluid dairy products. Appetite. 1988;10:37–44. - PubMed
-
- Takeda M, Sawano S, Imaizumi M, Fushiki T. Preference for corn oil in olfactory-blocked mice in the conditioned place preference test and the two-bottle choice test. Life Sci. 2001;69:847–854. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
