

Structure of the Fab fragment of F105, a broadly reactive anti-human immunodeficiency virus (HIV) antibody that recognizes the CD4 binding site of HIV type 1 gp120
- PMID: 16189008
- PMCID: PMC1235812
- DOI: 10.1128/JVI.79.20.13060-13069.2005
Structure of the Fab fragment of F105, a broadly reactive anti-human immunodeficiency virus (HIV) antibody that recognizes the CD4 binding site of HIV type 1 gp120
Abstract
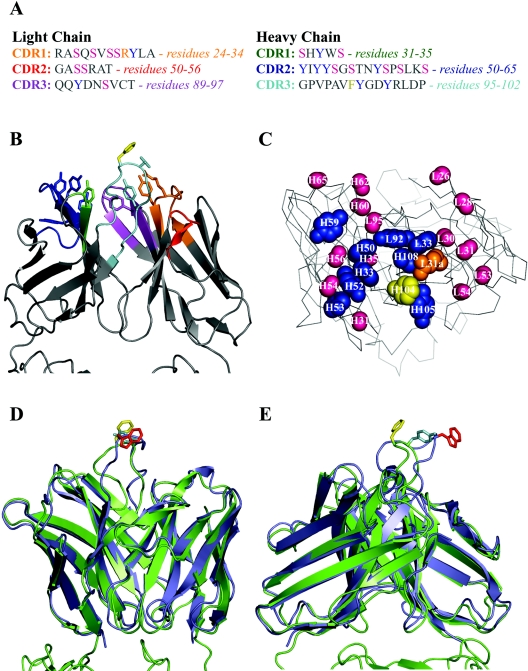
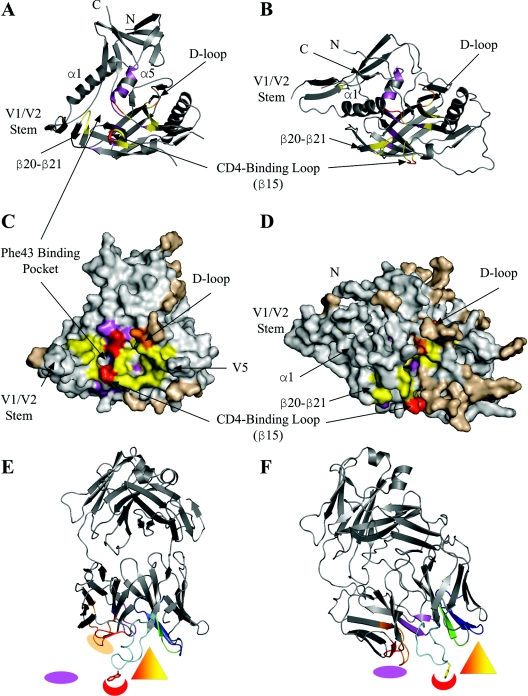
We have determined the crystal structure of the Fab fragment from F105, a broadly reactive human antibody with limited potency that recognizes the CD4 binding site of gp120. The structure reveals an extended CDR H3 loop with a phenylalanine residue at the apex and shows a striking pattern of serine and tyrosine residues. Modeling the interaction between gp120 and F105 suggests that the phenylalanine may recognize the binding pocket of gp120 used by Phe(43) of CD4 and that numerous tyrosine and serine residues form hydrogen bonds with the main chain atoms of gp120. A comparison of the F105 structure to that of immunoglobulin G1 b12, a much more potent and broadly neutralizing antibody with an overlapping epitope, suggests similarities that contribute to the broad recognition of human immunodeficiency virus by both antibodies. While the putative epitope for F105 shows significant overlap with that predicted for b12, it appears to differ from the b12 epitope in extending across the interface between the inner and outer domains of gp120. In contrast, the CDR loops of b12 appear to interact predominantly with the outer domain of gp120. The difference between the predicted epitopes for b12 and F105 suggests that the unique potency of b12 may arise from its ability to avoid the interface between the inner and outer domains of gp120.
Figures
Similar articles
-
Molecular features of the broadly neutralizing immunoglobulin G1 b12 required for recognition of human immunodeficiency virus type 1 gp120.J Virol. 2003 May;77(10):5863-76. doi: 10.1128/jvi.77.10.5863-5876.2003. J Virol. 2003. PMID: 12719580 Free PMC article.
-
Structural mimicry of CD4 by a cross-reactive HIV-1 neutralizing antibody with CDR-H2 and H3 containing unique motifs.J Mol Biol. 2006 Mar 17;357(1):82-99. doi: 10.1016/j.jmb.2005.12.062. Epub 2006 Jan 9. J Mol Biol. 2006. PMID: 16426633
-
Recognition properties of a panel of human recombinant Fab fragments to the CD4 binding site of gp120 that show differing abilities to neutralize human immunodeficiency virus type 1.J Virol. 1994 Aug;68(8):4821-8. doi: 10.1128/JVI.68.8.4821-4828.1994. J Virol. 1994. PMID: 7518527 Free PMC article.
-
Enhanced exposure of the CD4-binding site to neutralizing antibodies by structural design of a membrane-anchored human immunodeficiency virus type 1 gp120 domain.J Virol. 2009 May;83(10):5077-86. doi: 10.1128/JVI.02600-08. Epub 2009 Mar 4. J Virol. 2009. PMID: 19264769 Free PMC article.
-
Structural Features of Broadly Neutralizing Antibodies and Rational Design of Vaccine.Adv Exp Med Biol. 2018;1075:73-95. doi: 10.1007/978-981-13-0484-2_4. Adv Exp Med Biol. 2018. PMID: 30030790 Review.
Cited by
-
Comprehensive optimization of a single-chain variable domain antibody fragment as a targeting ligand for a cytotoxic nanoparticle.MAbs. 2015;7(1):42-52. doi: 10.4161/19420862.2014.985933. MAbs. 2015. PMID: 25484041 Free PMC article.
-
In vivo alteration of humoral responses to HIV-1 envelope glycoprotein gp120 by antibodies to the CD4-binding site of gp120.Virology. 2008 Mar 15;372(2):409-20. doi: 10.1016/j.virol.2007.10.044. Epub 2007 Dec 4. Virology. 2008. PMID: 18054978 Free PMC article.
-
Crystal structure of the neutralizing Llama V(HH) D7 and its mode of HIV-1 gp120 interaction.PLoS One. 2010 May 5;5(5):e10482. doi: 10.1371/journal.pone.0010482. PLoS One. 2010. PMID: 20463957 Free PMC article.
-
The W100 pocket on HIV-1 gp120 penetrated by b12 is not a target for other CD4bs monoclonal antibodies.Retrovirology. 2012 Jan 27;9:9. doi: 10.1186/1742-4690-9-9. Retrovirology. 2012. PMID: 22284192 Free PMC article.
-
Llama antibody fragments recognizing various epitopes of the CD4bs neutralize a broad range of HIV-1 subtypes A, B and C.PLoS One. 2012;7(3):e33298. doi: 10.1371/journal.pone.0033298. Epub 2012 Mar 15. PLoS One. 2012. PMID: 22438910 Free PMC article.
References
-
- Baba, T. W., V. Liska, R. Hofmann-Lehmann, J. Vlasak, W. Xu, S. Ayehunie, L. A. Cavacini, M. R. Posner, H. Katinger, G. Stiegler, B. J. Bernacky, T. A. Rizvi, R. Schmidt, L. R. Hill, M. E. Keeling, Y. Lu, J. E. Wright, T. C. Chou, and R. M. Ruprecht. 2000. Human neutralizing monoclonal antibodies of the IgG1 subtype protect against mucosal simian-human immunodeficiency virus infection. Nat. Med. 6:200-206. - PubMed
-
- Bagley, J., P. J. Dillon, C. Rosen, J. Robinson, J. Sodroski, and W. A. Marasco. 1994. Structural characterization of broadly neutralizing human monoclonal antibodies against the CD4 binding site of HIV-1 gp120. Mol. Immunol. 31:1149-1160. - PubMed
-
- Burton, D. R., R. C. Desrosiers, R. W. Doms, W. C. Koff, P. D. Kwong, J. P. Moore, G. J. Nabel, J. Sodroski, I. A. Wilson, and R. T. Wyatt. 2004. HIV vaccine design and the neutralizing antibody problem. Nat. Immunol. 5:233-236. - PubMed
-
- Cavacini, L. A., C. L. Emes, A. V. Wisnewski, J. Power, G. Lewis, D. Montefiori, and M. R. Posner. 1998. Functional and molecular characterization of human monoclonal antibody reactive with the immunodominant region of HIV type 1 glycoprotein 41. AIDS Res. Hum. Retrovir. 14:1271-1280. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
