

A role for auxin redistribution in the responses of the root system architecture to phosphate starvation in Arabidopsis
- PMID: 16040660
- PMCID: PMC1183395
- DOI: 10.1104/pp.105.060061
A role for auxin redistribution in the responses of the root system architecture to phosphate starvation in Arabidopsis
Abstract
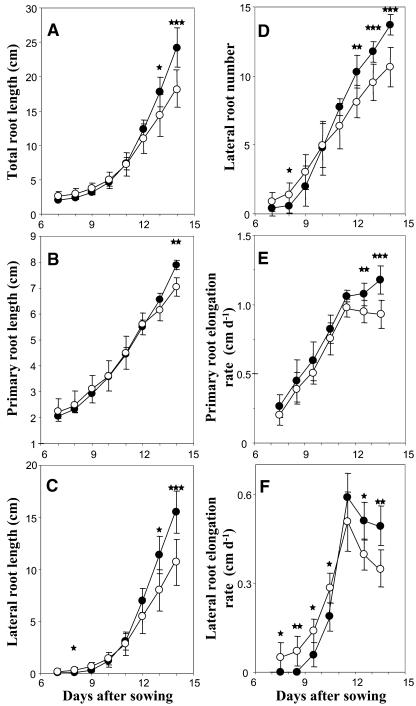
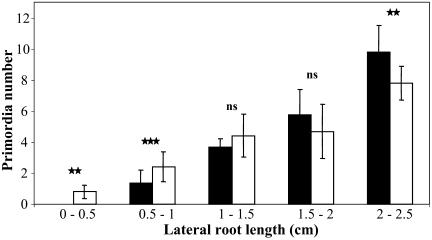
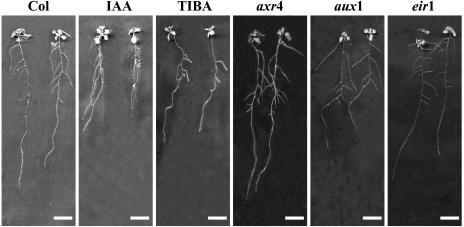
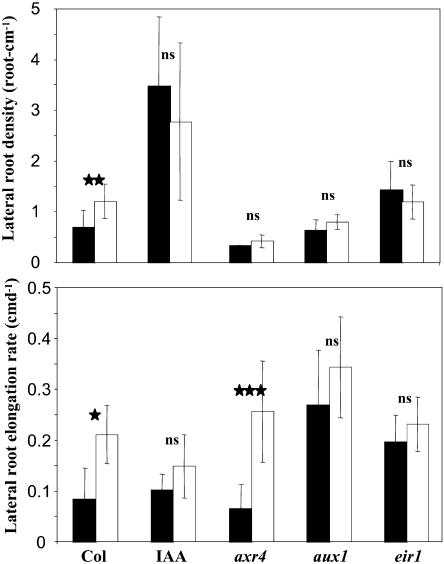
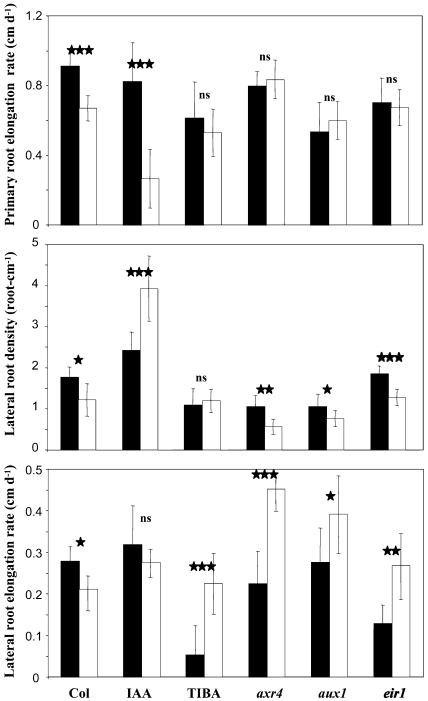
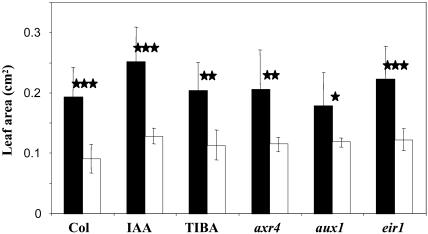
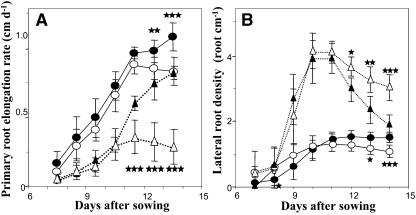
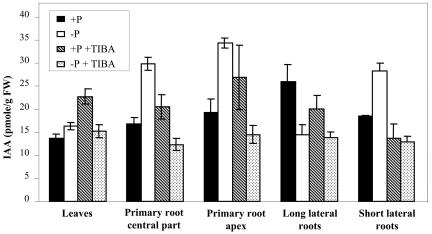
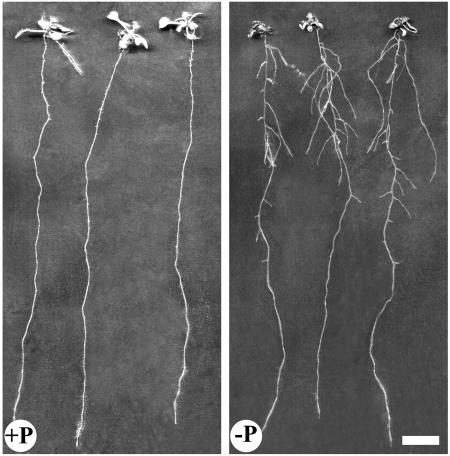
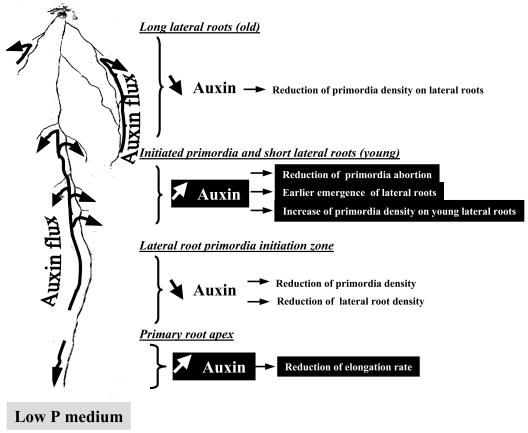
The changes in root system architecture (RSA) triggered by phosphate (P) deprivation were studied in Arabidopsis (Arabidopsis thaliana) plants grown for 14 d on 1 mM or 3 microM P. Two different temporal phases were observed in the response of RSA to low P. First, lateral root (LR) development was promoted between days 7 and 11 after germination, but, after day 11, all root growth parameters were negatively affected, leading to a general reduction of primary root (PR) and LR lengths and of LR density. Low P availability had contrasting effects on various stages of LR development, with a marked inhibition of primordia initiation but a strong stimulation of activation of the initiated primordia. The involvement of auxin signaling in these morphological changes was investigated in wild-type plants treated with indole-3-acetic acid or 2,3,5-triiodobenzoic acid and in axr4-1, aux1-7, and eir1-1 mutants. Most effects of low P on RSA were dramatically modified in the mutants or hormone-treated wild-type plants. This shows that auxin plays a major role in the P starvation-induced changes of root development. From these data, we hypothesize that several aspects of the RSA response to low P are triggered by local modifications of auxin concentration. A model is proposed that postulates that P starvation results in (1) an overaccumulation of auxin in the apex of the PR and in young LRs, (2) an overaccumulation of auxin or a change in sensitivity to auxin in the lateral primordia, and (3) a decrease in auxin concentration in the lateral primordia initiation zone of the PR and in old laterals. Measurements of local changes in auxin concentrations induced by low P, either by direct quantification or by biosensor expression pattern (DR5::beta-glucuronidase reporter gene), are in line with these hypotheses. Furthermore, the observation that low P availability mimicked the action of auxin in promoting LR development in the alf3 mutant confirmed that P starvation stimulates primordia emergence through increased accumulation of auxin or change in sensitivity to auxin in the primordia. Both the strong effect of 2,3,5-triiodobenzoic acid and the phenotype of the auxin-transport mutants (aux1, eir1) suggest that low P availability modifies local auxin concentrations within the root system through changes in auxin transport rather than auxin synthesis.
Figures


Similar articles
-
SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation.Plant Physiol. 2011 Feb;155(2):1000-12. doi: 10.1104/pp.110.165191. Epub 2010 Dec 14. Plant Physiol. 2011. PMID: 21156857 Free PMC article.
-
Differential effects of sucrose and auxin on localized phosphate deficiency-induced modulation of different traits of root system architecture in Arabidopsis.Plant Physiol. 2007 May;144(1):232-47. doi: 10.1104/pp.106.092130. Epub 2007 Mar 16. Plant Physiol. 2007. PMID: 17369438 Free PMC article.
-
Auxin and ethylene are involved in the responses of root system architecture to low boron supply in Arabidopsis seedlings.Physiol Plant. 2011 Jun;142(2):170-8. doi: 10.1111/j.1399-3054.2011.01459.x. Epub 2011 Mar 16. Physiol Plant. 2011. PMID: 21338369
-
A central role for the nitrate transporter NRT2.1 in the integrated morphological and physiological responses of the root system to nitrogen limitation in Arabidopsis.Plant Physiol. 2006 Mar;140(3):909-21. doi: 10.1104/pp.105.075721. Epub 2006 Jan 13. Plant Physiol. 2006. PMID: 16415211 Free PMC article.
-
The Plasticity of Root Systems in Response to External Phosphate.Int J Mol Sci. 2020 Aug 19;21(17):5955. doi: 10.3390/ijms21175955. Int J Mol Sci. 2020. PMID: 32824996 Free PMC article. Review.
Cited by
-
Modeling Auxin Signaling in Roots: Auxin Computations.Cold Spring Harb Perspect Biol. 2022 Feb 1;14(2):a040089. doi: 10.1101/cshperspect.a040089. Cold Spring Harb Perspect Biol. 2022. PMID: 34001532 Free PMC article. Review.
-
Phosphorus-Use-Efficiency Gene Identification in Fabaceae and RSL2 Expansion in Lupinus albus Is Associated with Low-Phosphorus Adaptation.Genes (Basel). 2024 Aug 9;15(8):1049. doi: 10.3390/genes15081049. Genes (Basel). 2024. PMID: 39202409 Free PMC article.
-
Natural genetic variation shapes root system responses to phytohormones in Arabidopsis.Plant J. 2018 Oct;96(2):468-481. doi: 10.1111/tpj.14034. Epub 2018 Sep 10. Plant J. 2018. PMID: 30030851 Free PMC article.
-
Arabidopsis Pht1;5 mobilizes phosphate between source and sink organs and influences the interaction between phosphate homeostasis and ethylene signaling.Plant Physiol. 2011 Jul;156(3):1149-63. doi: 10.1104/pp.111.174805. Epub 2011 May 31. Plant Physiol. 2011. PMID: 21628630 Free PMC article.
-
A scanner system for high-resolution quantification of variation in root growth dynamics of Brassica rapa genotypes.J Exp Bot. 2014 May;65(8):2039-48. doi: 10.1093/jxb/eru048. Epub 2014 Mar 6. J Exp Bot. 2014. PMID: 24604732 Free PMC article.
References
-
- Al-Ghazi Y, Muller B, Pinloche S, Tranbarger TJ, Nacry P, Rossignol M, Tardieu F, Doumas P (2003) Temporal response of Arabidopsis root architecture to phosphate starvation: evidence for the involvement of auxin signalling. Plant Cell Environ 26: 1053–1066
-
- Bates TR, Lynch JP (1996) Stimulation of root hair elongation in Arabidopsis thaliana by low phosphorus availability. Plant Cell Environ 19: 529–538
-
- Benkova E, Michniewicz M, Sauer M, Teichmann T, Seifertova D, Jurgens G, Friml J (2003) Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115: 591–602 - PubMed
-
- Bhalerao RP, Eklof J, Ljung K, Marchant A, Bennett M, Sandberg G (2002) Shoot-derived auxin is essential for early lateral root emergence in Arabidopsis seedlings. Plant J 29: 325–332 - PubMed
-
- Blakely LM, Durham M, Evans TA, Blakely RM (1982) Experimental studies on lateral root formation in radish seedlings roots. 1. General methods, developmental stages and spontaneous formation of laterals. Bot Gaz 143: 341–352
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
