

doi: 10.1128/JVI.79.13.8655-8660.2005.
Epstein-Barr virus (EBV) latent membrane protein 2A regulates B-cell receptor-induced apoptosis and EBV reactivation through tyrosine phosphorylation
Affiliations
- PMID: 15956608
- PMCID: PMC1143726
- DOI: 10.1128/JVI.79.13.8655-8660.2005
Item in Clipboard
Epstein-Barr virus (EBV) latent membrane protein 2A regulates B-cell receptor-induced apoptosis and EBV reactivation through tyrosine phosphorylation
J Virol.
2005 Jul.
Abstract
Epstein-Barr virus (EBV) is a human herpesvirus that establishes a lifelong latent infection of B cells. Within the immune system, apoptosis is a central mechanism in normal lymphocyte homeostasis both during early lymphocyte development and in response to antigenic stimuli. In this study, we found that latent membrane protein 2A (LMP2A) inhibited B-cell receptor (BCR)-induced apoptosis in Burkitt's lymphoma cell lines. Genistein, a specific inhibitor of tyrosine-specific protein kinases, blocked BCR-induced apoptosis and EBV reactivation in the cells. These findings indicate that LMP2A blocks BCR-induced cell apoptosis and EBV reactivation through the inhibition of activation of tyrosine kinases by BCR cross-linking.
Figures
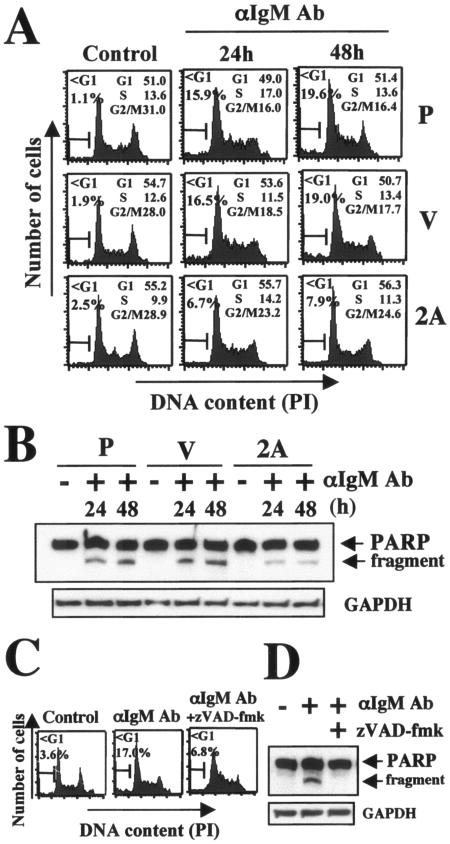
LMP2A inhibits BCR-induced DNA fragmentation and cleavage of PARP in Ramos cells. (A) Cells were seeded at 3 × 105 cells/ml, and cells were treated without (control) or with 35 μg/ml anti-IgM antibody (αIgAb) for 24 h or 48 h. Cells were analyzed for DNA content by propidium iodide (PI) staining and flow cytometry. Gates employed to ascertain cell cycle distribution and the percentage of cells with a sub-G1 (<G1) and G2/M DNA content are shown. These data are representative of three experiments. (B) PARP cleavage was analyzed by immunoblotting with a specific anti-PARP antibody. The full-length 113-kDa and 89-kDa cleaved PARP proteins are indicated. The amount of protein loaded in each lane was assessed by rehybridization of the filter with a specific antibody for human GAPDH. P, parental; V, vector control; 2A, LMP2A expressing. (C) DNA fragmentation and (D) evaluation of cleavage of PARP. Cells (3 × 105/ml) were preincubated for 1 h with or without zVAD-fmk (50 μM), and cells were then treated with 35 μg/ml anti-IgM antibody. After 24 h of incubation, cell cycles and PARP cleavage were analyzed as described in the legend to panel B. The amount of protein loaded in each lane was assessed by rehybridization of the filter with a specific antibody for human GAPDH.
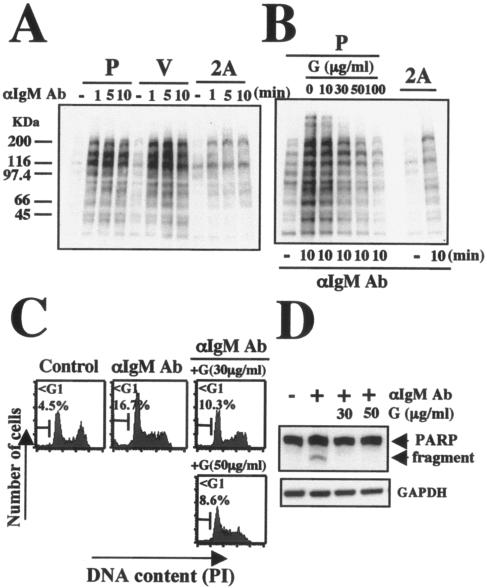
LMP2A or genistein inhibits BCR-induced tyrosine phosphorylation, DNA fragmentation, and cleavage PARP in Ramos cells. (A) Parental (P), vector control (V), and LMP2A-expressing (2A) Ramos cells (1 × 106/ml) were treated without (control [−]) or in the presence of 35 μg/ml anti-IgM antibody (αIgM Ab) for the indicated times (1, 5, or 10 min). (B) Cells (1 × 106/ml) were preincubated for 30 min with various concentrations of genistein, and then cells were treated with 35 μg/ml anti-IgM antibody for 10 min. Equal amounts of protein from the respective cells were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The levels of expression of phosphorylated tyrosine were determined by immunoblotting. (C) DNA fragmentation and (D) evaluation of cleavage of PARP. Cells (3 × 105/ml) were preincubated for 30 min with or without genistein, and cells were then treated with 35 μg/ml anti-IgM antibody. After 24 h of incubation, cell cycles and PARP cleavage were analyzed as described in the legend to Fig. 1. The amount of protein loaded in each lane was assessed by rehybridization of the filter with a specific antibody for human GAPDH.
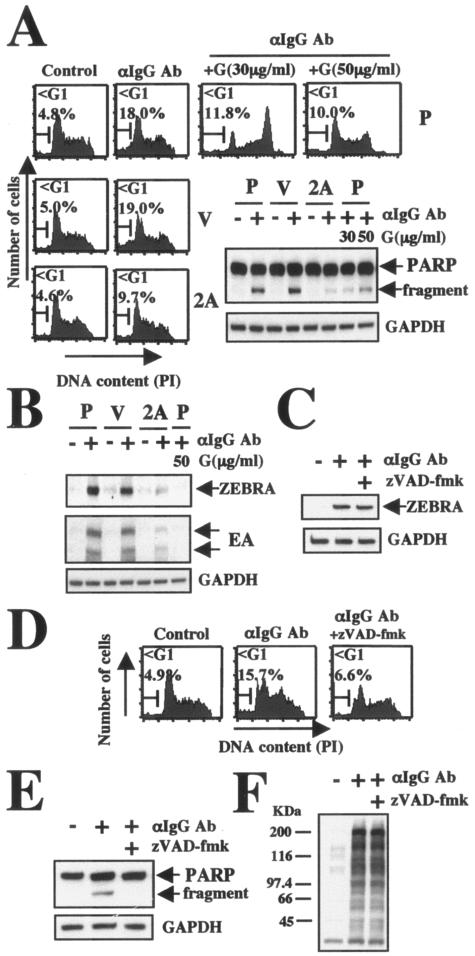
LMP2A or genistein blocks BCR-induced DNA fragmentation, cleavage of PARP, and EBV reactivation. (A) DNA fragmentation and evaluation of cleavage of PARP. Parental (P), vector control (V), and LMP2A-expressing (2A) Akata cells (5 × 105/ml) were preincubated for 30 min with or without genistein, and cells were then treated with 40 μg/ml anti-IgG antibody (αIgG Ab). After 24 h of incubation, cell cycle analyses and PARP cleavage were analyzed as described in the legend to Fig. 1. (B) EBV reactivation. Cells (5 × 105/ml) were preincubated for 30 min with or without genistein (G; 50 μg/ml), and cells were then treated with 40 μg/ml anti-IgG antibody. After 24 h of incubation, the expression of ZEBRA (38 kDa) and EA (52 kDa and 55 kDa) was analyzed by immunoblotting with a specific anti-ZEBRA antibody or with human EBV serum reactive with EBV early antigens. The amount of protein loaded in each lane was assessed by rehybridization of the filter with a specific antibody for human GAPDH. (C) EBV reactivation, (D) DNA fragmentation, and (E) cleavage of PARP with zVAD-fmk treatment. Cells (5 × 105/ml) were preincubated for 1 h with or without zVAD-fmk (50 μM), and cells were then treated with 40 μg/ml anti-IgG antibody. After 24 h of incubation, cell cycles and the expression of ZEBRA were analyzed as described in the legends to Fig. 1 and panel B. The amount of proteinloaded in each lane was assessed by rehybridization of the filter with a specific antibody for human GAPDH. (F) Tyrosine phosphorylation with zVAD-fmk treatment. Cells (1 × 106/ml) were preincubated for 1 h with or without zVAD-fmk (50 μM) followed by treatment with 35 μg/ml anti-IgM antibody for 10 min. Equal amounts of protein from the respective cells were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The levels of expression of phosphorylated tyrosine was determined by immunoblotting.
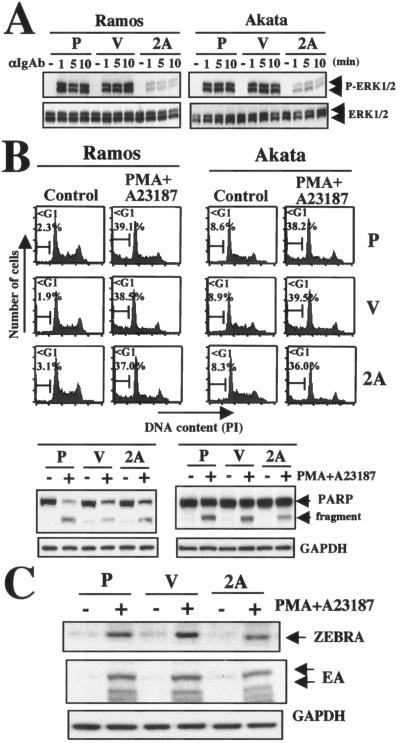
PMA and calcium ionophore A23187 induce apoptosis and EBV reactivation in LMP2A-expressing Ramos and Akata cells. (A) Parental (P), vector control (V), and LMP2A-expressing (2A) Ramos and Akata cells (1 × 106/ml) were treated without (control [−]) or with of 35 μg/ml anti-IgM antibody or 40 μg/ml anti-IgG antibody for the indicated times (1, 5, or 10 min). Equal amounts of protein from the respective cells were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. ERK1/2 phosphorylation was detected with anti-phospho MAPK antibody. Upper and lower arrows indicate ERK1 and ERK2, respectively. The lower panels show equal loading of proteins and the expression of total ERK (ERK1/2). (B) DNA fragmentation, evaluation of cleavage of PARP, and (C) EBV reactivation. Parental (P), vector control (V), and LMP2A (2A)-expressing Ramos (3 × 105/ml) and Akata (5 × 105/ml) cells were treated with 20 ng/ml PMA and 7.5 μM A23187. After 24 h of incubation, cell cycles, PARP cleavage, and the expression of ZEBRA and EA were analyzed as described in the legends to Fig. 1 and Fig. 3.
Similar articles
-
Epstein-Barr virus Latent Membrane Protein 2A (LMP2A)-mediated changes in Fas expression and Fas-dependent apoptosis: Role of Lyn/Syk activation.Cell Immunol. 2015 Oct;297(2):108-19. doi: 10.1016/j.cellimm.2015.08.001. Epub 2015 Aug 4. Cell Immunol. 2015. PMID: 26255694 Free PMC article.
-
Epstein-Barr virus coopts lipid rafts to block the signaling and antigen transport functions of the BCR.Immunity. 2001 Jan;14(1):57-67. doi: 10.1016/s1074-7613(01)00089-9. Immunity. 2001. PMID: 11163230
-
Tyrosines 60, 64, and 101 of Epstein-Barr virus LMP2A are not essential for blocking B cell signal transduction.Virology. 1999 Oct 25;263(2):485-95. doi: 10.1006/viro.1999.9964. Virology. 1999. PMID: 10544120
-
Epstein-Barr virus protein LMP2A regulates reactivation from latency by negatively regulating tyrosine kinases involved in sIg-mediated signal transduction.Infect Agents Dis. 1994 Apr-Jun;3(2-3):128-36. Infect Agents Dis. 1994. PMID: 7812651 Review.
-
The effects of the Epstein-Barr virus latent membrane protein 2A on B cell function.Int Rev Immunol. 2001;20(6):805-35. doi: 10.3109/08830180109045591. Int Rev Immunol. 2001. PMID: 11913951 Review.
Cited by
-
Epstein-Barr virus LMP2A imposes sensitivity to apoptosis.J Gen Virol. 2010 Sep;91(Pt 9):2197-202. doi: 10.1099/vir.0.021444-0. Epub 2010 May 19. J Gen Virol. 2010. PMID: 20484564 Free PMC article.
-
K1 and K15 of Kaposi's Sarcoma-Associated Herpesvirus Are Partial Functional Homologues of Latent Membrane Protein 2A of Epstein-Barr Virus.J Virol. 2015 Jul;89(14):7248-61. doi: 10.1128/JVI.00839-15. Epub 2015 May 6. J Virol. 2015. PMID: 25948739 Free PMC article.
-
Epstein-Barr virus latent membrane protein 2A mediates transformation through constitutive activation of the Ras/PI3-K/Akt Pathway.J Virol. 2007 Sep;81(17):9299-306. doi: 10.1128/JVI.00537-07. Epub 2007 Jun 20. J Virol. 2007. PMID: 17582000 Free PMC article.
-
Epstein-Barr virus-encoded LMP2A induces an epithelial-mesenchymal transition and increases the number of side population stem-like cancer cells in nasopharyngeal carcinoma.PLoS Pathog. 2010 Jun 3;6(6):e1000940. doi: 10.1371/journal.ppat.1000940. PLoS Pathog. 2010. PMID: 20532215 Free PMC article.
-
Epstein-Barr virus in Burkitt's lymphoma: a role for latent membrane protein 2A.Cell Cycle. 2010 Mar 1;9(5):901-8. doi: 10.4161/cc.9.5.10840. Epub 2010 Mar 3. Cell Cycle. 2010. PMID: 20160479 Free PMC article.
References
-
- An, S., and K. A. Knox. 1996. Ligation of CD40 rescues Ramos-Burkitt lymphoma B cells from calcium ionophore- and antigen receptor-triggered apoptosis by inhibiting activation of the cysteine protease CPP32/Yama and cleavage of its substrate PARP. FEBS Lett. 386:115-122. - PubMed
-
- Babcock, G. J., L. L. Decker, M. Volk, and D. A. Thorley-Lawson. 1998. EBV persistence in memory B cells in vivo. Immunity 9:395-404. - PubMed
-
- Babcock, G. J., D. Hochberg, and A. D. Thorley-Lawson. 2000. The expression pattern of Epstein-Barr virus latent genes in vivo is dependent upon the differentiation stage of the infected B cell. Immunity 13:497-506. - PubMed
-
- Barnes, S. 1995. Effect of genistein on in vitro and in vivo models of cancer. J. Nutr. 125:777-783. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
