

Apoptotic cells deliver processed antigen to dendritic cells for cross-presentation
- PMID: 15839733
- PMCID: PMC1084338
- DOI: 10.1371/journal.pbio.0030185
Apoptotic cells deliver processed antigen to dendritic cells for cross-presentation
Abstract
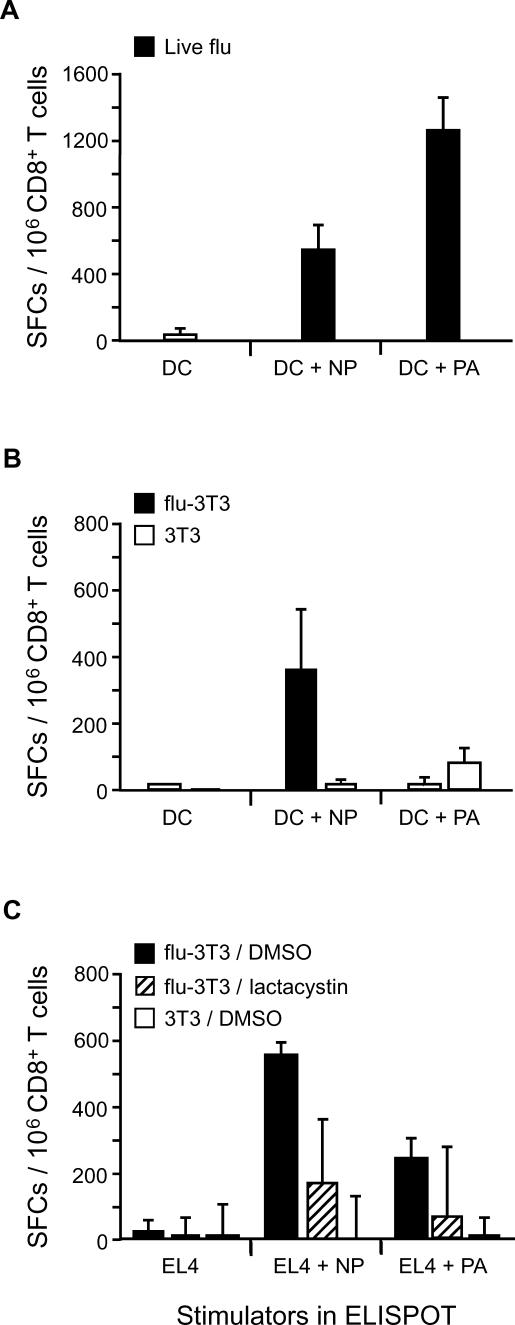
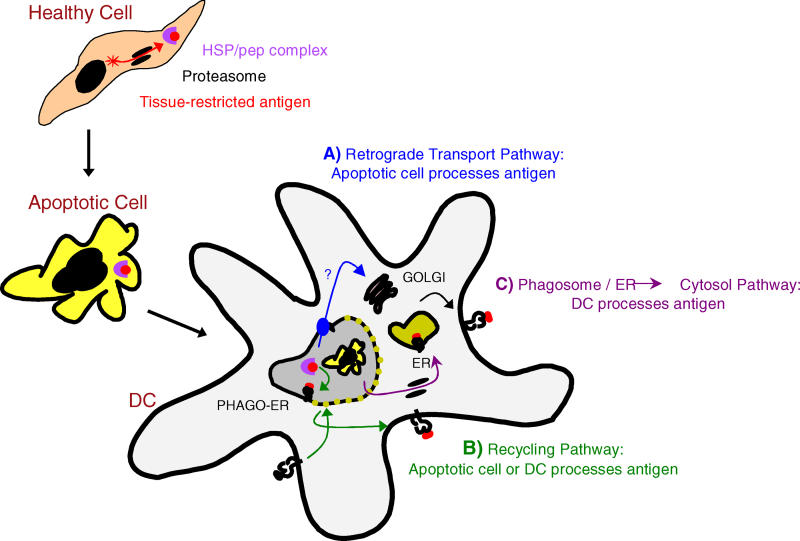
Antigen derived from engulfed apoptotic cells can be cross-presented by dendritic cells (DCs) for the generation of major histocompatibility class I/peptide complexes. While the early events of recognition and internalization of the dying cell have been characterized, the antigen-processing pathway or pathways remain unknown. We established a mouse model assaying for the activation of polyclonal T cells reactive to antigen derived from apoptotic cells, and demonstrated two distinct pathways for the trafficking of exogenous epitopes. In the first, exogenous antigen is dependent on the DC's expression of a functional transporter associated with antigen processing (TAP). Surprisingly, we found evidence that a second pathway exists in which transfer of processed antigen from the dying cell allows formation of major histocompatibility class I/peptide complexes in TAP-deficient DCs. In vivo data suggest that in situations of stress (e.g., viral infection), this latter pathway may be more efficient, illustrating that dying cells may preselect immunologically important antigenic determinants.
Figures




Similar articles
-
Constitutive macropinocytosis allows TAP-dependent major histocompatibility complex class I presentation of exogenous soluble antigen by bone marrow-derived dendritic cells.Eur J Immunol. 1997 Jan;27(1):280-8. doi: 10.1002/eji.1830270141. Eur J Immunol. 1997. PMID: 9022030
-
Cross-presentation of virus-like particles by skin-derived CD8(-) dendritic cells: a dispensable role for TAP.Eur J Immunol. 2002 Mar;32(3):818-25. doi: 10.1002/1521-4141(200203)32:3<818::AID-IMMU818>3.0.CO;2-U. Eur J Immunol. 2002. PMID: 11870626
-
Control of cross-presentation during dendritic cell maturation.Eur J Immunol. 2004 Feb;34(2):398-407. doi: 10.1002/eji.200324508. Eur J Immunol. 2004. PMID: 14768044
-
Bacterial antigen delivery systems: phagocytic processing of bacterial antigens for MHC-I and MHC-II presentation to T cells.Behring Inst Mitt. 1997 Feb;(98):197-211. Behring Inst Mitt. 1997. PMID: 9382741 Review.
-
The ABCs of immunology: structure and function of TAP, the transporter associated with antigen processing.Physiology (Bethesda). 2004 Aug;19:216-24. doi: 10.1152/physiol.00002.2004. Physiology (Bethesda). 2004. PMID: 15304636 Review.
Cited by
-
Human macrophages and dendritic cells can equally present MART-1 antigen to CD8(+) T cells after phagocytosis of gamma-irradiated melanoma cells.PLoS One. 2012;7(7):e40311. doi: 10.1371/journal.pone.0040311. Epub 2012 Jul 2. PLoS One. 2012. PMID: 22768350 Free PMC article.
-
Immunogenic Apoptosis as a Novel Tool for Anticancer Vaccine Development.Int J Mol Sci. 2018 Feb 16;19(2):594. doi: 10.3390/ijms19020594. Int J Mol Sci. 2018. PMID: 29462947 Free PMC article. Review.
-
Zinc supplementation is required for the cytotoxic and immunogenic effects of chemotherapy in chemoresistant p53-functionally deficient cells.Oncoimmunology. 2013 Sep 1;2(9):e26198. doi: 10.4161/onci.26198. Epub 2013 Sep 12. Oncoimmunology. 2013. PMID: 24228232 Free PMC article.
-
Spatiotemporal Regulation of Hsp90-Ligand Complex Leads to Immune Activation.Front Immunol. 2016 May 24;7:201. doi: 10.3389/fimmu.2016.00201. eCollection 2016. Front Immunol. 2016. PMID: 27252703 Free PMC article. Review.
-
Poly (I:C) enhances the anti-tumor activity of canine parvovirus NS1 protein by inducing a potent anti-tumor immune response.Tumour Biol. 2016 Sep;37(9):12089-12102. doi: 10.1007/s13277-016-5093-z. Epub 2016 May 21. Tumour Biol. 2016. PMID: 27209409
References
-
- Savill J. Recognition and phagocytosis of cells undergoing apoptosis. Br Med Bull. 1997;53:491–508. - PubMed
-
- Guermonprez P, Valladeau J, Zitvogel L, Thery C, Amigorena S. Antigen presentation and T cell stimulation by dendritic cells. Annu Rev Immunol. 2002;20:621–667. - PubMed
-
- Heath WR, Carbone FR. Cross-presentation in viral immunity and self-tolerance. Nat Rev Immunol. 2001;1:126–134. - PubMed
-
- Albert ML, Darnell JC, Bender A, Francisco LM, Bhardwaj N, et al. Tumor-specific killer cells in paraneoplastic cerebellar degeneration. Nat Med. 1998;4:1321–1324. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
