

Interaction between the CD8 coreceptor and major histocompatibility complex class I stabilizes T cell receptor-antigen complexes at the cell surface
- PMID: 15837791
- PMCID: PMC2441837
- DOI: 10.1074/jbc.M500555200
Interaction between the CD8 coreceptor and major histocompatibility complex class I stabilizes T cell receptor-antigen complexes at the cell surface
Abstract
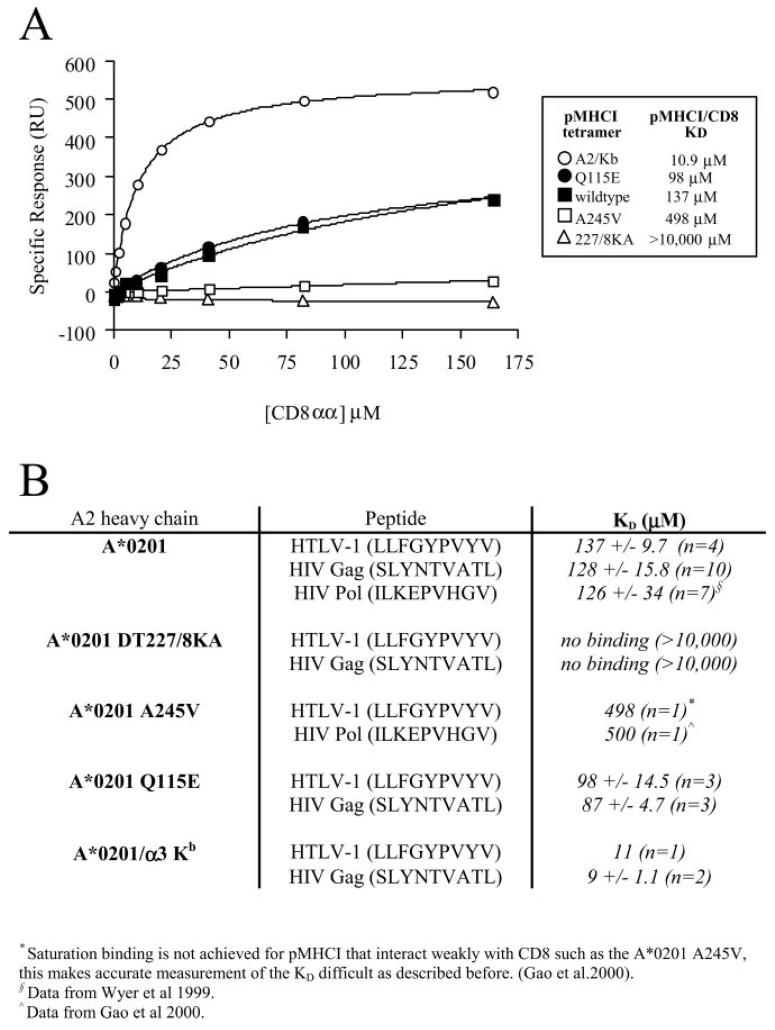
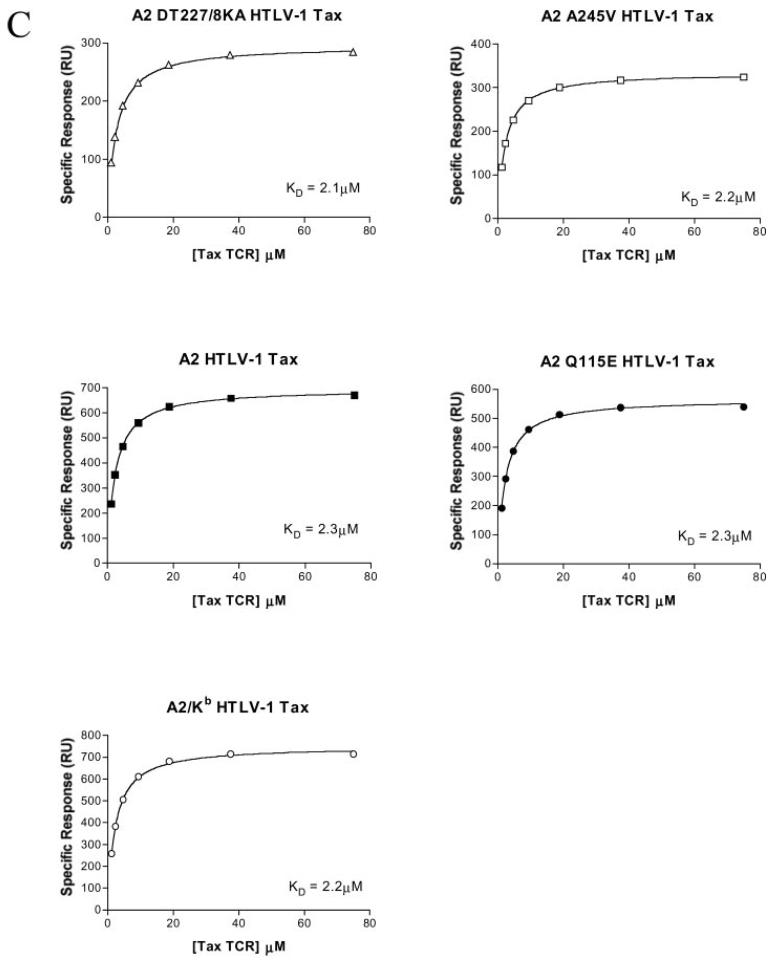
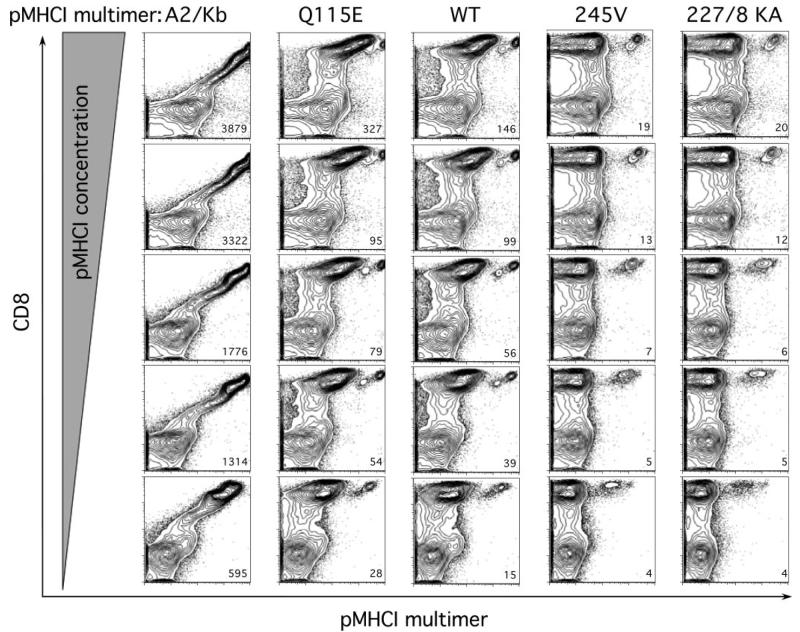
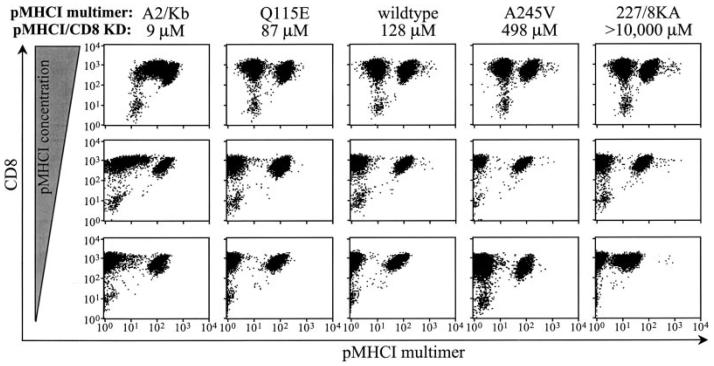
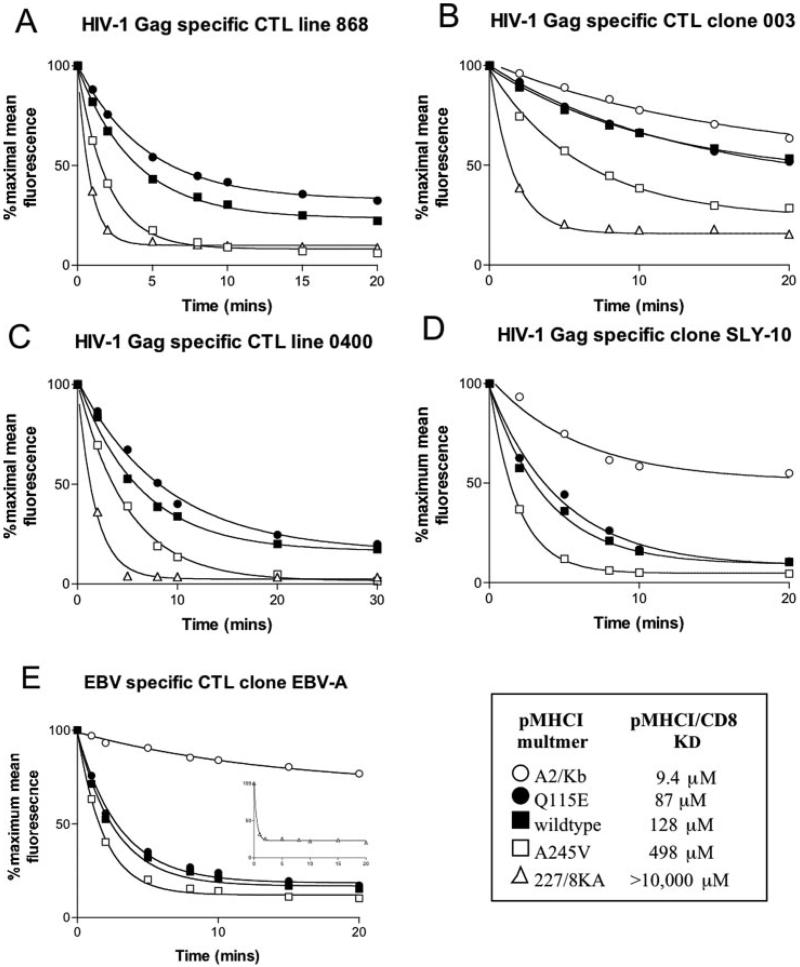
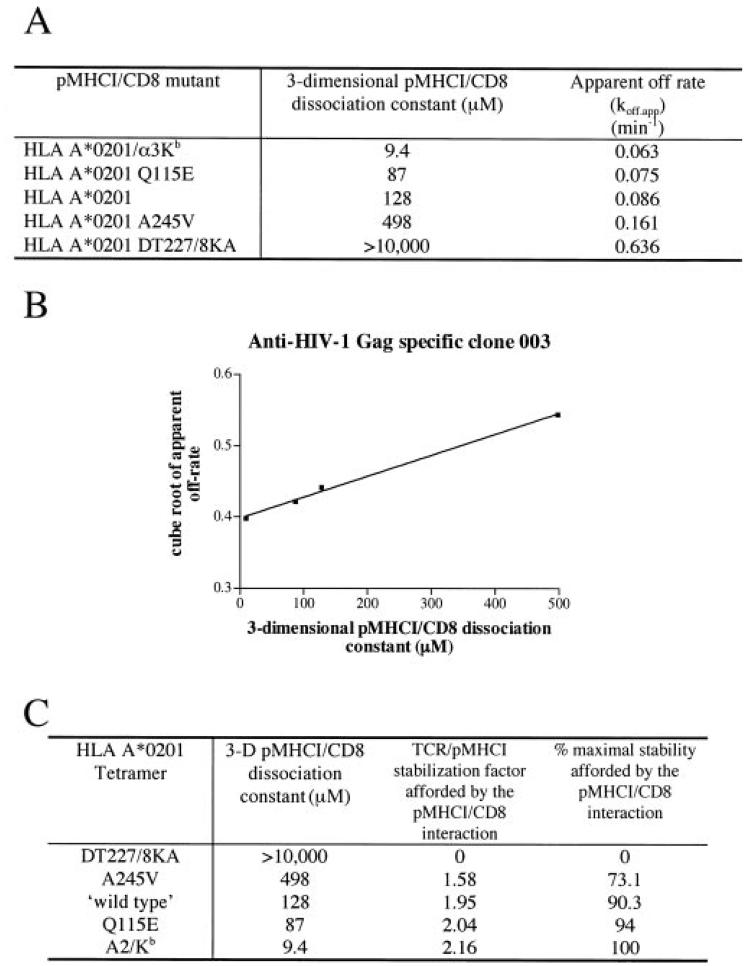
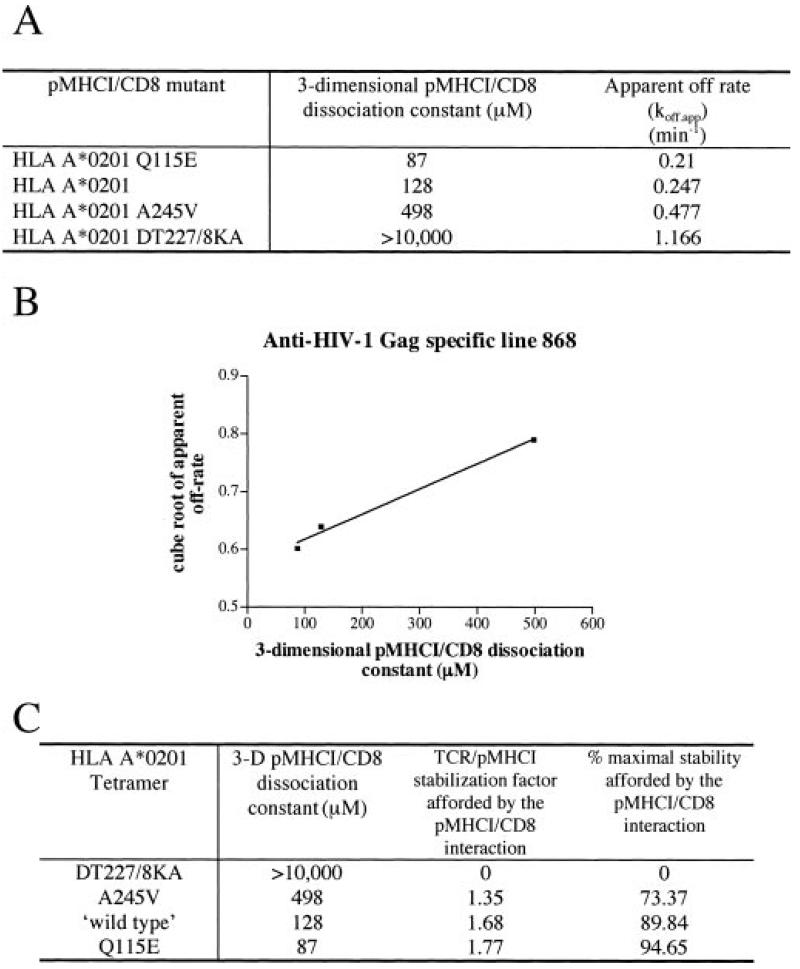
The off-rate (k(off)) of the T cell receptor (TCR)/peptide-major histocompatibility complex class I (pMHCI) interaction, and hence its half-life, is the principal kinetic feature that determines the biological outcome of TCR ligation. However, it is unclear whether the CD8 coreceptor, which binds pMHCI at a distinct site, influences this parameter. Although biophysical studies with soluble proteins show that TCR and CD8 do not bind cooperatively to pMHCI, accumulating evidence suggests that TCR associates with CD8 on the T cell surface. Here, we titrated and quantified the contribution of CD8 to TCR/pMHCI dissociation in membrane-constrained interactions using a panel of engineered pMHCI mutants that retain faithful TCR interactions but exhibit a spectrum of affinities for CD8 of >1,000-fold. Data modeling generates a "stabilization factor" that preferentially increases the predicted TCR triggering rate for low affinity pMHCI ligands, thereby suggesting an important role for CD8 in the phenomenon of T cell cross-reactivity.
Figures
Similar articles
-
Different T cell receptor affinity thresholds and CD8 coreceptor dependence govern cytotoxic T lymphocyte activation and tetramer binding properties.J Biol Chem. 2007 Aug 17;282(33):23799-810. doi: 10.1074/jbc.M700976200. Epub 2007 May 31. J Biol Chem. 2007. PMID: 17540778
-
CD8 coreceptor-mediated focusing can reorder the agonist hierarchy of peptide ligands recognized via the T cell receptor.Proc Natl Acad Sci U S A. 2021 Jul 20;118(29):e2019639118. doi: 10.1073/pnas.2019639118. Proc Natl Acad Sci U S A. 2021. PMID: 34272276 Free PMC article.
-
T cell receptor engagement of peptide-major histocompatibility complex class I does not modify CD8 binding.Mol Immunol. 2008 May;45(9):2700-9. doi: 10.1016/j.molimm.2007.12.009. Epub 2008 Feb 19. Mol Immunol. 2008. PMID: 18243322
-
Interplay between the TCR/CD3 complex and CD4 or CD8 in the activation of cytotoxic T lymphocytes.Immunol Rev. 1989 Jun;109:119-41. doi: 10.1111/j.1600-065x.1989.tb00022.x. Immunol Rev. 1989. PMID: 2527803 Review.
-
Molecular coordination of alphabeta T-cell receptors and coreceptors CD8 and CD4 in their recognition of peptide-MHC ligands.Trends Immunol. 2002 Aug;23(8):408-13. doi: 10.1016/s1471-4906(02)02282-2. Trends Immunol. 2002. PMID: 12133804 Review.
Cited by
-
Affinity-matured HLA class II dimers for robust staining of antigen-specific CD4+ T cells.Nat Biotechnol. 2021 Aug;39(8):958-967. doi: 10.1038/s41587-021-00836-4. Epub 2021 Mar 1. Nat Biotechnol. 2021. PMID: 33649568
-
Modulation of MHC binding by lateral association of TCR and coreceptor.Biophys J. 2012 Nov 7;103(9):1890-8. doi: 10.1016/j.bpj.2012.09.011. Biophys J. 2012. PMID: 23199917 Free PMC article.
-
IL-21 can supplement suboptimal Lck-independent MAPK activation in a STAT-3-dependent manner in human CD8(+) T cells.J Immunol. 2012 Feb 15;188(4):1609-19. doi: 10.4049/jimmunol.1003446. Epub 2012 Jan 11. J Immunol. 2012. PMID: 22238455 Free PMC article.
-
Structure, function, and immunomodulation of the CD8 co-receptor.Front Immunol. 2024 Aug 26;15:1412513. doi: 10.3389/fimmu.2024.1412513. eCollection 2024. Front Immunol. 2024. PMID: 39253084 Free PMC article. Review.
-
CD8+ T-cell specificity is compromised at a defined MHCI/CD8 affinity threshold.Immunol Cell Biol. 2017 Jan;95(1):68-76. doi: 10.1038/icb.2016.85. Epub 2016 Nov 8. Immunol Cell Biol. 2017. PMID: 27670790 Free PMC article.
References
-
- Davis MM, Boniface JJ, Reich Z, Lyons D, Hampl J, Arden B, Chien Y. Annu. Rev. Immunol. 1998;16:523–544. - PubMed
-
- Rudolph MG, Wilson IA. Curr. Opin. Immunol. 2002;14:52–65. - PubMed
-
- Gao GF, Tormo J, Gerth UC, Wyer JR, McMichael AJ, Stuart DI, Bell JI, Jones EY, Jakobsen BK. Nature. 1997;387:630–634. - PubMed
-
- Kern PS, Teng MK, Smolyar A, Liu JH, Liu J, Hussey RE, Spoerl R, Chang HC, Reinherz EL, Wang JH. Immunity. 1998;9:519–530. - PubMed
-
- Janeway CA., Jr. Annu. Rev. Immunol. 1992;10:645–674. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
