

Chromatin and RNAi factors protect the C. elegans germline against repetitive sequences
- PMID: 15774721
- PMCID: PMC1074315
- DOI: 10.1101/gad.332305
Chromatin and RNAi factors protect the C. elegans germline against repetitive sequences
Abstract
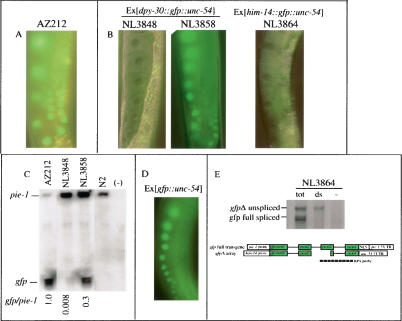
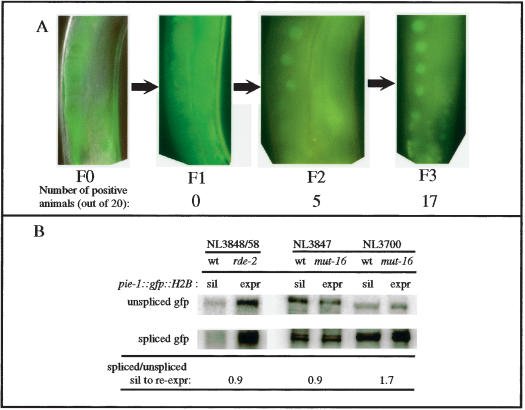
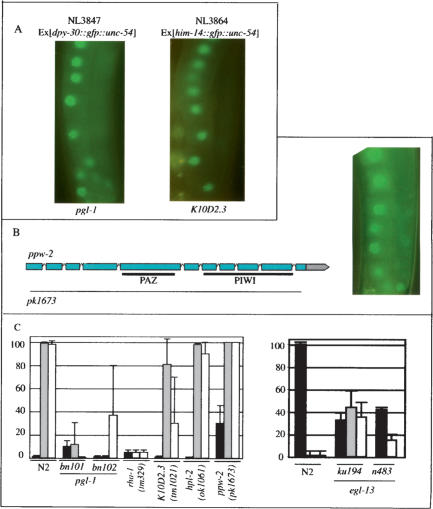
Protection of genomes against invasion by repetitive sequences, such as transposons, viruses, and repetitive transgenes, involves strong and selective silencing of these sequences. During silencing of repetitive transgenes, a trans effect ("cosuppression") occurs that results in silencing of cognate endogenous genes. Here we report RNA interference (RNAi) screens performed to catalog genes required for cosuppression in the Caenorhabditis elegans germline. We find factors with a putative role in chromatin remodeling and factors involved in RNAi. Together with molecular data also presented in this study, these results suggest that in C. elegans repetitive sequences trigger transcriptional gene silencing using RNAi and chromatin factors.
Figures
Similar articles
-
Transgene-mediated cosuppression and RNA interference enhance germ-line apoptosis in Caenorhabditis elegans.Proc Natl Acad Sci U S A. 2012 Feb 28;109(9):3440-5. doi: 10.1073/pnas.1107390109. Epub 2012 Feb 13. Proc Natl Acad Sci U S A. 2012. PMID: 22331911 Free PMC article.
-
Transcriptional silencing of a transgene by RNAi in the soma of C. elegans.Genes Dev. 2005 Mar 15;19(6):683-96. doi: 10.1101/gad.1247705. Epub 2005 Mar 1. Genes Dev. 2005. PMID: 15741313 Free PMC article.
-
Decoupling the downstream effects of germline nuclear RNAi reveals that H3K9me3 is dispensable for heritable RNAi and the maintenance of endogenous siRNA-mediated transcriptional silencing in Caenorhabditis elegans.Epigenetics Chromatin. 2017 Feb 15;10:6. doi: 10.1186/s13072-017-0114-8. eCollection 2017. Epigenetics Chromatin. 2017. PMID: 28228846 Free PMC article.
-
RNAi mechanisms in Caenorhabditis elegans.FEBS Lett. 2005 Oct 31;579(26):5932-9. doi: 10.1016/j.febslet.2005.08.001. Epub 2005 Aug 9. FEBS Lett. 2005. PMID: 16162338 Review.
-
Multigenerational Regulation of the Caenorhabditis elegans Chromatin Landscape by Germline Small RNAs.Annu Rev Genet. 2019 Dec 3;53:289-311. doi: 10.1146/annurev-genet-112618-043505. Epub 2019 May 31. Annu Rev Genet. 2019. PMID: 31150586 Free PMC article. Review.
Cited by
-
Germ granules and gene regulation in the Caenorhabditis elegans germline.Genetics. 2022 Mar 3;220(3):iyab195. doi: 10.1093/genetics/iyab195. Genetics. 2022. PMID: 35239965 Free PMC article. Review.
-
Germ Granules Govern Small RNA Inheritance.Curr Biol. 2019 Sep 9;29(17):2880-2891.e4. doi: 10.1016/j.cub.2019.07.054. Epub 2019 Aug 1. Curr Biol. 2019. PMID: 31378614 Free PMC article.
-
Transgenerational functions of small RNA pathways in controlling gene expression in C. elegans.Epigenetics. 2014 Jan;9(1):37-44. doi: 10.4161/epi.26795. Epub 2013 Oct 25. Epigenetics. 2014. PMID: 24162759 Free PMC article. Review.
-
Formation of artificial chromosomes in Caenorhabditis elegans and analyses of their segregation in mitosis, DNA sequence composition and holocentromere organization.Nucleic Acids Res. 2021 Sep 20;49(16):9174-9193. doi: 10.1093/nar/gkab690. Nucleic Acids Res. 2021. PMID: 34417622 Free PMC article.
-
Caenorhabditis elegans SynMuv B gene activity is down-regulated during a viral infection to enhance RNA interference.bioRxiv [Preprint]. 2024 Jul 16:2024.07.12.603258. doi: 10.1101/2024.07.12.603258. bioRxiv. 2024. PMID: 39071373 Free PMC article. Preprint.
References
-
- Bernstein E., Caudy, A.A., Hammond, S.M., and Hannon, G.J. 2001. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409: 363–366. - PubMed
-
- Chen C., Simard, M.J., Tabara, H., Brownell, D.R., McCollough, J.A., and Mello, C.C. 2005. A member of the polymerase β nucleotidyltransferase superfamily is required for RNA interference in C. elegans. Curr. Biol. 15: 378–383. - PubMed
-
- Denli A.M., Tops, B.B., Plasterk, R.H., Ketting, R.F., and Hannon, G.J. 2004. Processing of primary microRNAs by the Microprocessor complex. Nature 432: 231–235. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials