

Transcriptional silencing of a transgene by RNAi in the soma of C. elegans
- PMID: 15741313
- PMCID: PMC1065722
- DOI: 10.1101/gad.1247705
Transcriptional silencing of a transgene by RNAi in the soma of C. elegans
Abstract
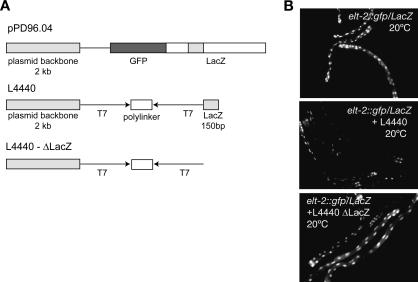
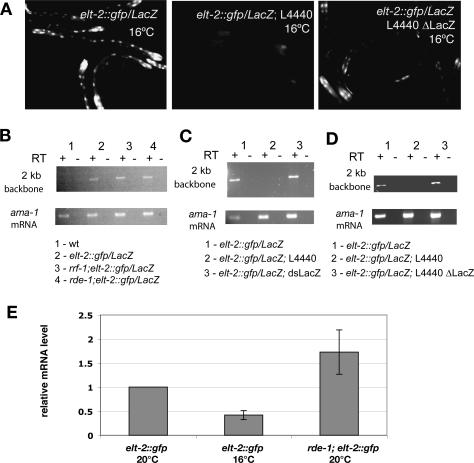
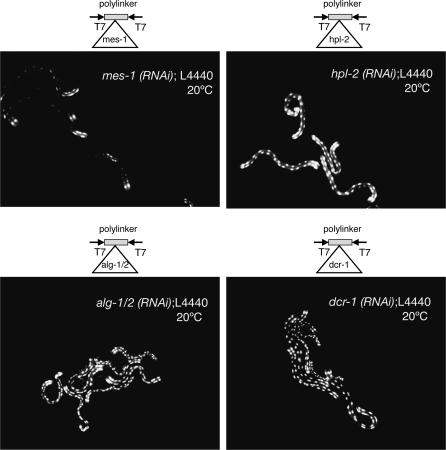
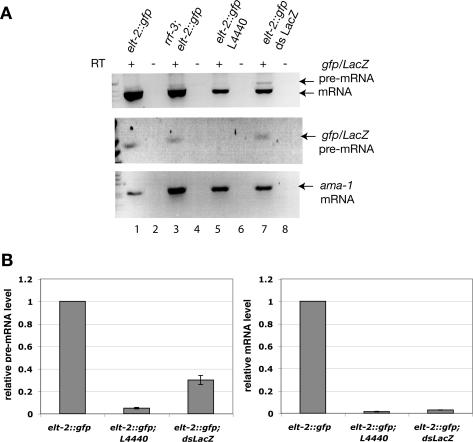
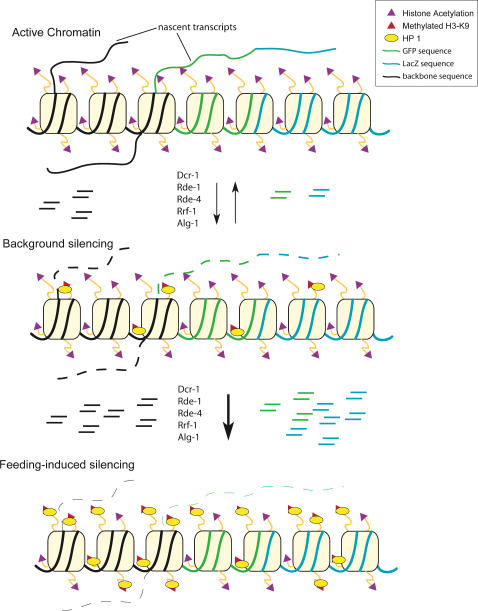
The silencing of transgene expression at the level of transcription in the soma of Caenorhabditis elegans through an RNAi-dependent pathway has not been previously characterized. Most gene silencing due to RNAi in C. elegans occurs at the post-transcriptional level. We observed transcriptional silencing when worms containing the elt-2::gfp/LacZ transgene were fed RNA produced from the commonly used L4440 vector. The transgene and the vector share plasmid backbone sequences. This transgene silencing depends on multiple RNAi pathway genes, including dcr-1, rde-1, rde-4, and rrf-1. Unlike post-transcriptional gene silencing in worms, elt-2::gfp/LacZ silencing is dependent on the PAZ-PIWI protein Alg-1 and on the HP1 homolog Hpl-2. The latter is a chromatin silencing factor, and expression of the transgene is inhibited at the level of intron-containing precursor mRNA. This inhibition is accompanied by a decrease in the acetylation of histones associated with the transgene. This transcriptional silencing in the soma can be distinguished from transgene silencing in the germline by its inability to be transmitted across generations and its dependence on the rde-1 gene. We therefore define this type of silencing as RNAi-induced Transcriptional Gene Silencing (RNAi-TGS). Additional chromatin-modifying components affecting RNAi-TGS were identified in a candidate RNAi screen.
Figures


Similar articles
-
In C. elegans, high levels of dsRNA allow RNAi in the absence of RDE-4.PLoS One. 2008;3(12):e4052. doi: 10.1371/journal.pone.0004052. Epub 2008 Dec 29. PLoS One. 2008. PMID: 19112503 Free PMC article.
-
The dsRNA binding protein RDE-4 interacts with RDE-1, DCR-1, and a DExH-box helicase to direct RNAi in C. elegans.Cell. 2002 Jun 28;109(7):861-71. doi: 10.1016/s0092-8674(02)00793-6. Cell. 2002. PMID: 12110183
-
Transgene-Assisted Genetic Screen Identifies rsd-6 and Novel Genes as Key Components of Antiviral RNA Interference in Caenorhabditis elegans.J Virol. 2018 Aug 16;92(17):e00416-18. doi: 10.1128/JVI.00416-18. Print 2018 Sep 1. J Virol. 2018. PMID: 29950414 Free PMC article.
-
RNAi mechanisms in Caenorhabditis elegans.FEBS Lett. 2005 Oct 31;579(26):5932-9. doi: 10.1016/j.febslet.2005.08.001. Epub 2005 Aug 9. FEBS Lett. 2005. PMID: 16162338 Review.
-
RNA interference: genetic wand and genetic watchdog.Nat Cell Biol. 2000 Feb;2(2):E31-6. doi: 10.1038/35000102. Nat Cell Biol. 2000. PMID: 10655601 Review.
Cited by
-
piRNAs initiate an epigenetic memory of nonself RNA in the C. elegans germline.Cell. 2012 Jul 6;150(1):65-77. doi: 10.1016/j.cell.2012.06.015. Epub 2012 Jun 25. Cell. 2012. PMID: 22738726 Free PMC article.
-
Current understanding on micro RNAs and its regulation in response to Mycobacterial infections.J Biomed Sci. 2013 Feb 28;20(1):14. doi: 10.1186/1423-0127-20-14. J Biomed Sci. 2013. PMID: 23448104 Free PMC article. Review.
-
Identification and characterization of two novel classes of small RNAs in the mouse germline: retrotransposon-derived siRNAs in oocytes and germline small RNAs in testes.Genes Dev. 2006 Jul 1;20(13):1732-43. doi: 10.1101/gad.1425706. Epub 2006 Jun 9. Genes Dev. 2006. PMID: 16766679 Free PMC article.
-
Germ granules and gene regulation in the Caenorhabditis elegans germline.Genetics. 2022 Mar 3;220(3):iyab195. doi: 10.1093/genetics/iyab195. Genetics. 2022. PMID: 35239965 Free PMC article. Review.
-
Piwi/PRG-1 Argonaute and TGF-β Mediate Transgenerational Learned Pathogenic Avoidance.Cell. 2019 Jun 13;177(7):1827-1841.e12. doi: 10.1016/j.cell.2019.05.024. Epub 2019 Jun 6. Cell. 2019. PMID: 31178117 Free PMC article.
References
-
- Bartel D.P. 2004. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 116: 281-297. - PubMed
-
- Bernstein E., Caudy, A.A., Hammond, S.M., and Hannon, G.J. 2001. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409: 363-366. - PubMed
-
- Caudy A.A., Ketting, R.F., Hammond, S.M., Denli, A.M., Bathoorn, A.M., Tops, B.B., Silva, J.M., Myers, M.M., Hannon, G.J., and Plasterk, R.H. 2003. A micrococcal nuclease homologue in RNAi effector complexes. Nature 425: 411-414. - PubMed
-
- Chan S.W., Zilberman, D., Xie, Z., Johansen, L.K., Carrington, J.C., and Jacobsen, S.E. 2004. RNA silencing genes control de novo DNA methylation. Science 303: 1336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials