

ARFGAP1 plays a central role in coupling COPI cargo sorting with vesicle formation
- PMID: 15657398
- PMCID: PMC2171589
- DOI: 10.1083/jcb.200404008
ARFGAP1 plays a central role in coupling COPI cargo sorting with vesicle formation
Abstract
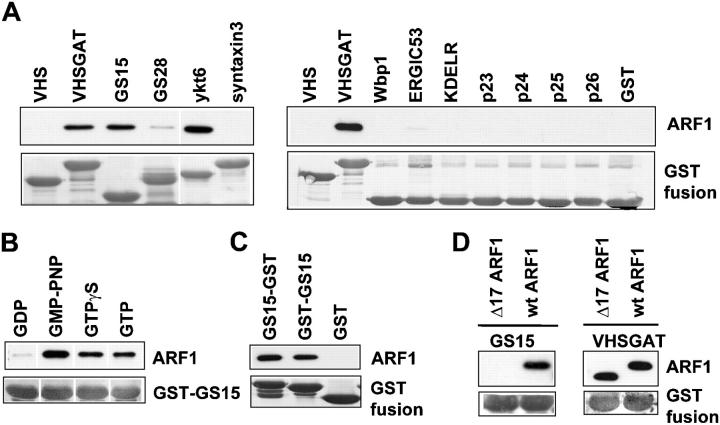
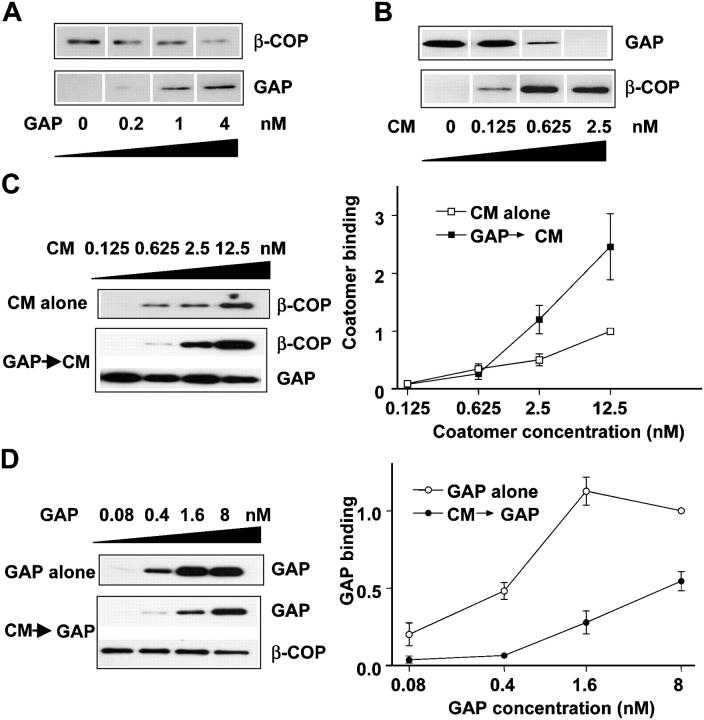
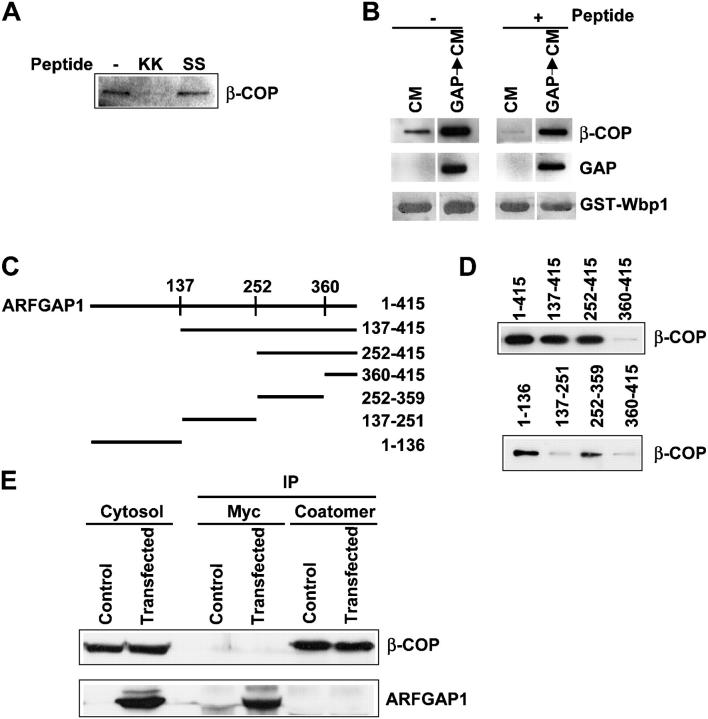
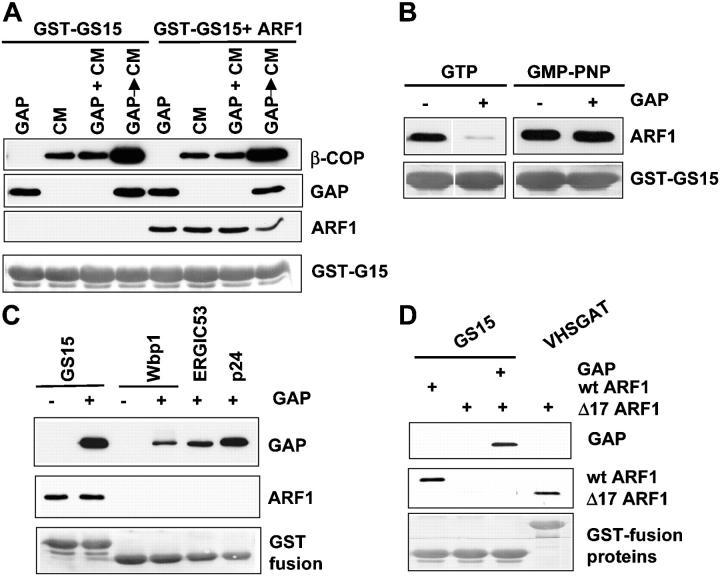
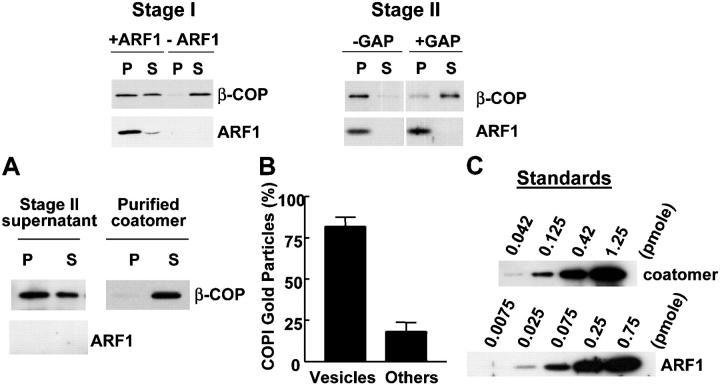
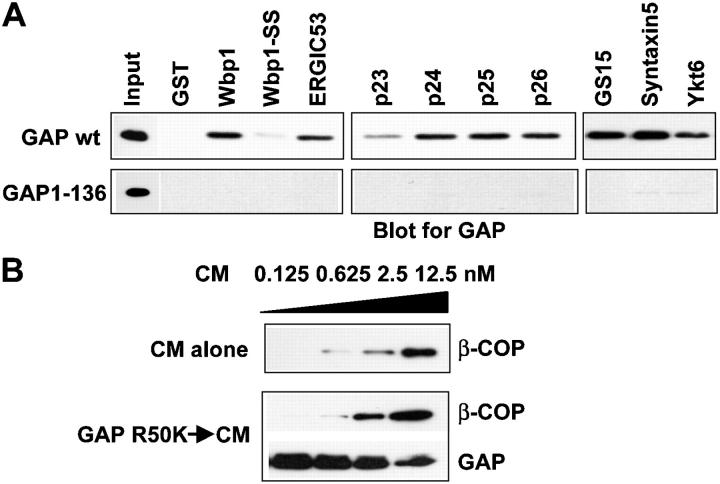
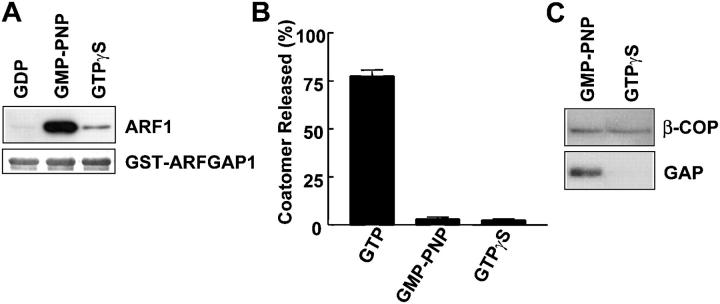
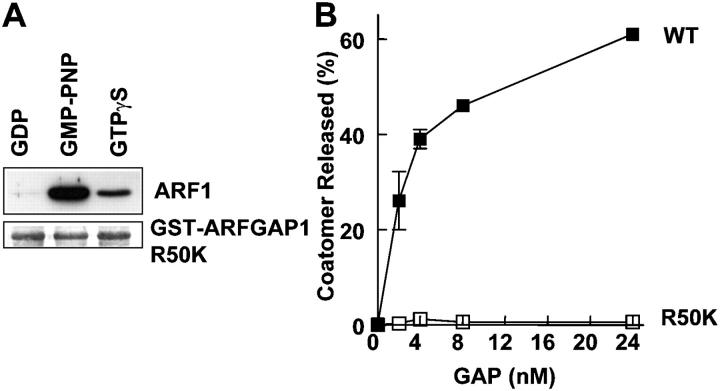
Examining how key components of coat protein I (COPI) transport participate in cargo sorting, we find that, instead of ADP ribosylation factor 1 (ARF1), its GTPase-activating protein (GAP) plays a direct role in promoting the binding of cargo proteins by coatomer (the core COPI complex). Activated ARF1 binds selectively to SNARE cargo proteins, with this binding likely to represent at least a mechanism by which activated ARF1 is stabilized on Golgi membrane to propagate its effector functions. We also find that the GAP catalytic activity plays a critical role in the formation of COPI vesicles from Golgi membrane, in contrast to the prevailing view that this activity antagonizes vesicle formation. Together, these findings indicate that GAP plays a central role in coupling cargo sorting and vesicle formation, with implications for simplifying models to describe how these two processes are coupled during COPI transport.
Figures
Similar articles
-
ArfGAP1 dynamics and its role in COPI coat assembly on Golgi membranes of living cells.J Cell Biol. 2005 Mar 28;168(7):1053-63. doi: 10.1083/jcb.200410142. J Cell Biol. 2005. PMID: 15795316 Free PMC article.
-
ARFGAP1 promotes the formation of COPI vesicles, suggesting function as a component of the coat.J Cell Biol. 2002 Oct 14;159(1):69-78. doi: 10.1083/jcb.200206015. Epub 2002 Oct 14. J Cell Biol. 2002. PMID: 12379802 Free PMC article.
-
Functional reconstitution of COPI coat assembly and disassembly using chemically defined components.Proc Natl Acad Sci U S A. 2003 Jul 8;100(14):8253-7. doi: 10.1073/pnas.1432391100. Epub 2003 Jun 27. Proc Natl Acad Sci U S A. 2003. PMID: 12832619 Free PMC article.
-
The COPI system: molecular mechanisms and function.FEBS Lett. 2009 Sep 3;583(17):2701-9. doi: 10.1016/j.febslet.2009.07.032. Epub 2009 Jul 22. FEBS Lett. 2009. PMID: 19631211 Review.
-
Membrane curvature and the control of GTP hydrolysis in Arf1 during COPI vesicle formation.Biochem Soc Trans. 2005 Aug;33(Pt 4):619-22. doi: 10.1042/BST0330619. Biochem Soc Trans. 2005. PMID: 16042557 Review.
Cited by
-
Reconstitution of COPI Vesicle and Tubule Formation.Methods Mol Biol. 2016;1496:63-74. doi: 10.1007/978-1-4939-6463-5_6. Methods Mol Biol. 2016. PMID: 27632002 Free PMC article.
-
Ubiquitylation of epsilon-COP by PIRH2 and regulation of the secretion of PSA.Mol Cell Biochem. 2008 Jan;307(1-2):73-82. doi: 10.1007/s11010-007-9586-3. Epub 2007 Aug 25. Mol Cell Biochem. 2008. PMID: 17721809
-
Cellular interactome analysis of vaccinia virus K7 protein identifies three transport machineries as binding partners for K7.Virus Genes. 2017 Dec;53(6):814-822. doi: 10.1007/s11262-017-1504-5. Epub 2017 Aug 16. Virus Genes. 2017. PMID: 28815417
-
ArfGAP1 inhibits mTORC1 lysosomal localization and activation.EMBO J. 2021 Jun 15;40(12):e106412. doi: 10.15252/embj.2020106412. Epub 2021 May 14. EMBO J. 2021. PMID: 33988249 Free PMC article.
-
Conserved functions of membrane active GTPases in coated vesicle formation.Science. 2009 Sep 4;325(5945):1217-20. doi: 10.1126/science.1171004. Science. 2009. PMID: 19729648 Free PMC article. Review.
References
-
- Bigay, J., P. Gounon, S. Robineau, and B. Antonny. 2003. Lipid packing sensed by ArfGAP1 couples COPI coat disassembly to membrane bilayer curvature. Nature. 426:563–566. - PubMed
-
- Bonifacino, J.S., and B.S. Glick. 2004. The mechanisms of vesicle budding and fusion. Cell. 116:153–166. - PubMed
-
- Cosson, P., and F. Letourneur. 1994. Coatomer interaction with di-lysine endoplasmic reticulum retention motifs. Science. 263:1629–1631. - PubMed
-
- Cukierman, E., I. Huber, M. Rotman, and D. Cassel. 1995. The ARF1-GTPase-activating protein: zinc finger motif and Golgi complex localization. Science. 270:1999–2002. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
