

Review
doi: 10.1128/MMBR.68.4.745-770.2004.
Metabolic interdependence of obligate intracellular bacteria and their insect hosts
Affiliations
- PMID: 15590782
- PMCID: PMC539007
- DOI: 10.1128/MMBR.68.4.745-770.2004
Item in Clipboard
Review
Metabolic interdependence of obligate intracellular bacteria and their insect hosts
Microbiol Mol Biol Rev.
2004 Dec.
Abstract
Mutualistic associations of obligate intracellular bacteria and insects have attracted much interest in the past few years due to the evolutionary consequences for their genome structure. However, much less attention has been paid to the metabolic ramifications for these endosymbiotic microorganisms, which have to compete with but also to adapt to another metabolism--that of the host cell. This review attempts to provide insights into the complex physiological interactions and the evolution of metabolic pathways of several mutualistic bacteria of aphids, ants, and tsetse flies and their insect hosts.
Figures

Glycolysis, TCA cycle, and gluconeogenesis in the different endosymbiotic bacteria. In Buchnera and “Candidatus Blochmannia,” glucose is oxidized to acetyl-CoA, while in Wigglesworthia, the pathway works in the opposite, gluconeogenetic direction. In Buchnera the TCA cycle is reduced to α-ketoglutarate dehydrogenase activity only, while in “Candidatus Blochmannia” and Wigglesworthia, most energy-yielding steps are conserved. Transport systems for sugars and glutamate are indicated by colored circles or boxes. Features missing in the respective organism are highlighted in red. Steps between glycerol-3-phosphate and PEP are conserved in all endosymbionts and are not shown in the figure. Steps generating reductive power in the form of NADH or leading to ATP formation by substrate-level phosphorylation are indicated in green. αKG, α-ketoglutarate; Mqo, malate:quinone oxidoreductase; AspC, aspartate aminotransferase.
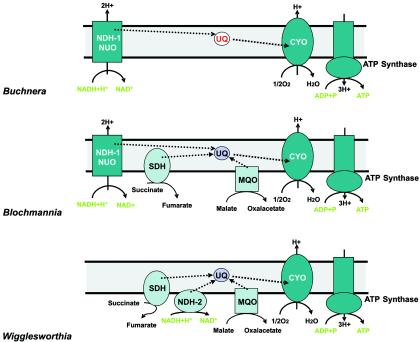
Electron transport chains of the endosymbiotic bacteria. In Buchnera, the electron transport chain consists merely of NADH dehydrogenase I (alternative designation, NUO) and cytochrome o oxidase (CYO). As indicated in red, ubiquinone (UQ) cannot be synthesized by Buchnera but has to be provided by the host. In “Candidatus Blochmannia” and Wigglesworthia, electron transport is more complex and succinate dehydrogenase (SDH) and malate:quinone oxidoreductase (MQO) are present. In Wigglesworthia, NDH-1 (NUO) is replaced by NDH-2, which does not translocate protons across the membrane. Oxidoreductases coupling electron transport with proton translocation are shown in dark blue, and oxidoreductases which are not coupling are shown in light blue.
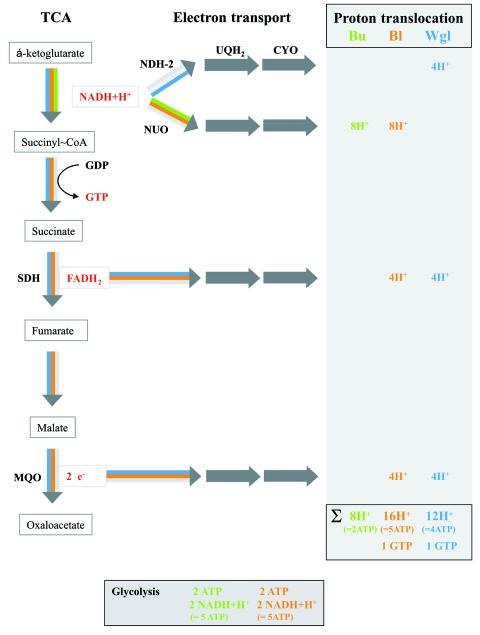
Energy yield and proton translocation. Buchnera, “Candidatus Blochmannia,” and Wigglesworthia are shown in the colors indicated in the graphic. The right panel summarizes proton translocation in the respective organisms. In Buchnera, a total of eight H+ ions are translocated, assuming that two H+/e− are translocated by the proton-pumping enzymes in the electron transport chain. In “Candidatus Blochmannia,” SDH and MQO add another four H+ ions each to the total sum. In Wigglesworthia, H+ translocation is reduced by four H+ ions compared with “Candidatus Blochmannia,” due to the non-proton-pumping NADH dehydrogenase NDH-2. ATP yield was counted on the assumption of three H+ ions per ATP. NDH-2, NADH ubiquinone oxidoreductase II; NUO, NADH ubiquinone oxidoreductase I (NDH-1); SDH, succinate dehydrogenase; MQO, malate:quinone oxidoreductase; CYO, cytochrome o oxidase.
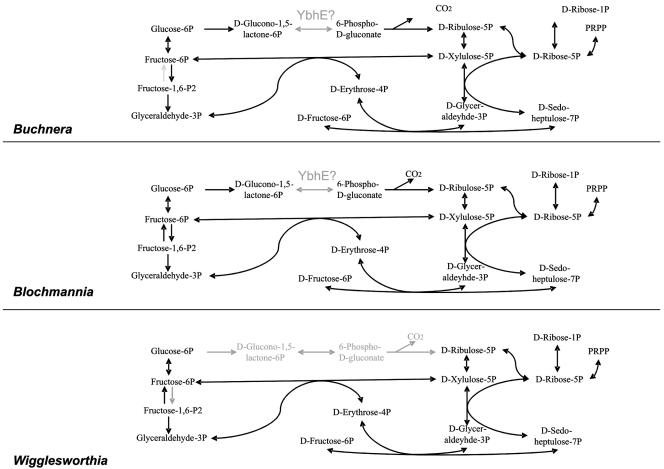
Pentose phosphate pathway. In Wigglesworthia, the oxidative branch of the pentose phosphate pathway is missing and only the regenerative steps are present. Missing steps are shown in grey. The YbhE protein is thought to catalyze the conversion of d -glucono-1,5-lactone-6-phosphate to 6-phospho-d -gluconate (see the text for details).
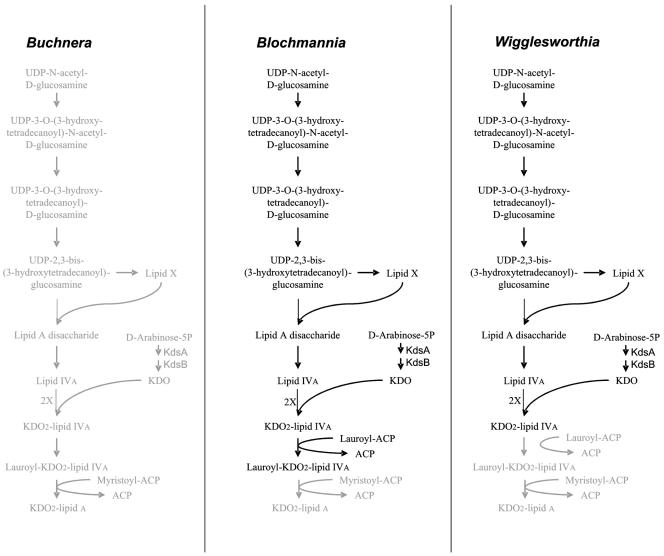
Lipid A biosynthesis. Biosynthesis pathways of lipid A in the different endosymbiotic bacteria are shown. Steps missing in the respective organisms are highlighted in grey; e.g., Buchnera is missing the entire pathway. In contrast, in “Candidatus Blochmannia” and Wigglesworthia, only the final steps involving acylation of KDO2-lipid IVa are partially missing.
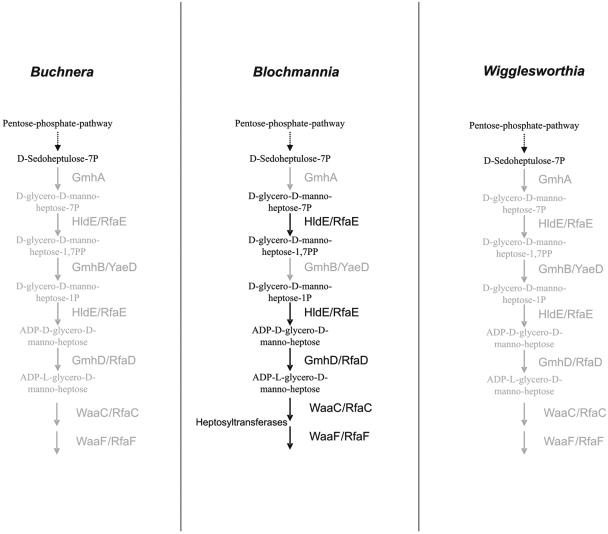
Modification of LPS by heptoses. In the free-living Enterobacteriaceae, the LPS core is further modified with heptoses. All three endosymbionts apparently are unable to synthesize the respective heptoses. The biosynthesis and modification enzymes were entirely (Buchnera and Wigglesworthia) or partially (“Candidatus Blochmannia”) lost, and missing steps are highlighted in grey.

Synthesis of amino sugars. “Candidatus Blochmannia” and Wigglesworthia are able to build up UDP-N-acetylmuramate from fructose-6-phosphate. In Buchnera, the transition of d -glucosamine-6-phosphate to UDP-N-acetylglucosamine seems to be blocked, and the missing steps are highlighted in grey. However, the lack of the respective enzymes very probably can be compensated for by provision of N-acetyl-d -glucosamine-1-phosphate by the host.
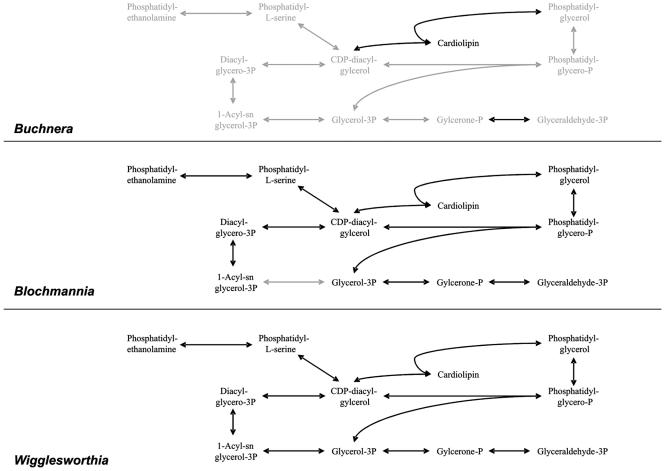
Phospholipid synthesis. In Buchnera the complete biosynthetic pathway except cardiolipin synthase is missing. If cardiolipin synthase is still active, the respective precursors, CDP-diacylglycerol and phosphatidylglycerol, have to be provided by the host. In “Candidatus Blochmannia,” only the glycerol-3-phosphate O-acyltransferase specific for the first acyltransfer is missing, while in Wigglesworthia the pathway is complete. Missing steps are highlighted in grey.
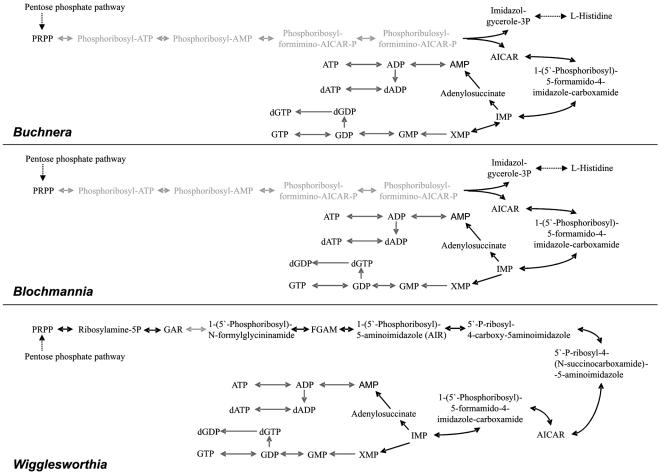
Purine biosynthesis. In Wigglesworthia, with the exception of the phosphoribosylglycinamide formyltransferase PurN, the complete purine biosynthetic pathway is present. In Buchnera and “Candidatus Blochmannia,” the first steps, leading from PRPP to AICAR, have been lost entirely. However, AICAR is also an intermediate of histidine biosynthesis in these two organisms, allowing purine biosynthesis by the combination of histidine and purine biosynthesis pathways. The conversion of purine intermediates “downstream” of the intermediates AMP, IMP, and XMP is not entirely clear from the data derived from the genome sequence, since some dedicated enzymes apparently are missing. However, as described in the text, the respective reactions are likely to be carried out by related enzymes, possibly as a result of an expansion of the substrate specificity of these enzymes. The light grey arrows highlight missing steps in purine biosynthesis, which are replaced by parts of the ehistidine biosynthesis pathuray in Blochmannia and Buchnera, and the dark grey arrows show the parts of the purine biosynthesis pathway which very probably can be carried out by the bacteria.

Pyrimidine biosynthesis. Buchnera is able to synthesize UMP, but most successive steps are missing, while “Candidatus Blochmannia” seems to need UMP as the starting material but, with the exception of trymidylate synthase (ThyA), can catalyze all consecutive steps. Only in Wigglesworthia is the pyrimidine biosynthetic pathway complete. Missing steps are highlighted in grey.
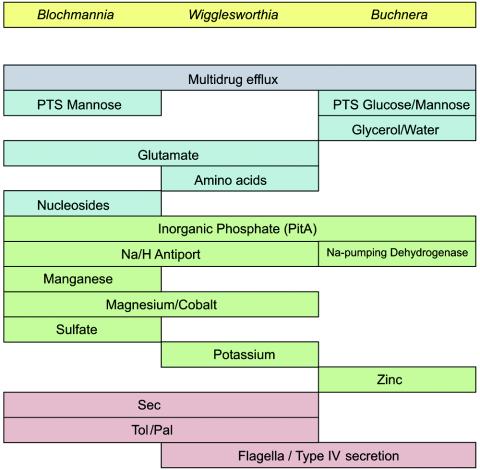
Overview of the transport capacities in the different endosymbionts. In most cases, only the general transport capabilities of the endosymbiotic bacteria are shown. The dedicated transport systems may differ among the various bacteria. The Buchnera genome encodes the smallest number of transport systems. Colors indicate multidrug transport systems (gray), metabolite transport systems (blue), ion transport systems (green), and macromolecule transport systems (red). For details see the text.

Models of pathway evolution in endosymbionts. Several theories of pathway evolution (left side) and their endosymbiontic modification (right side) are shown. Pathways may have evolved (bold arrows) in different ways: (a) de novo (all reactions evolved independently from an unknown origin), (b) backwards (retroevolution), (c) by specialization of a multifunctional enzyme (left side, with multiple substrates in blue), (d) duplication of the complete pathway, or (e) by recruitment of enzymes from different pathways. The scenarios shown on the right evolve with specific modifications in endosymbionts; selection pressure is sometimes in the opposite direction, pathway reductions occur to a large extent, and selected pathways evolve, e.g., to provide metabolites useful for the host.
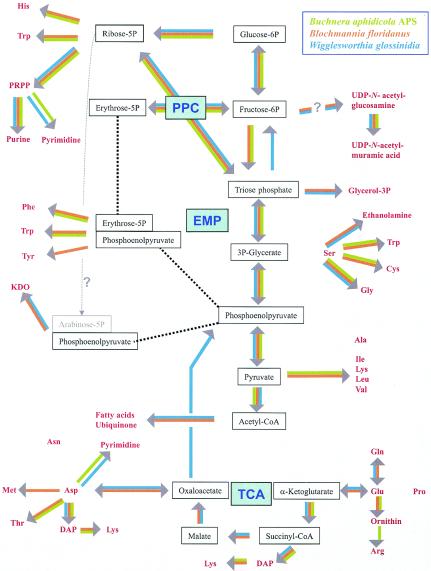
Overview and comparison of the central intermediate metabolism of the endosymbiotic bacteria. Building-block biosynthesis of the endosymbiotic bacteria “Candidatus Blochmannia” (orange), Buchnera (green), and Wigglesworthia (blue) is shown as deduced from their genomic sequences. Important intermediates or end products are shown in red. Gray arrows indicate lack of the respective pathways. EMP, Embden-Meyerhoff-Parnass pathway (glycolysis); PPC, pentose phosphate cycle.
Similar articles
-
Genome evolution in bacterial endosymbionts of insects.Nat Rev Genet. 2002 Nov;3(11):850-61. doi: 10.1038/nrg931. Nat Rev Genet. 2002. PMID: 12415315 Review.
-
Reductive evolution of bacterial genome in insect gut environment.Genome Biol Evol. 2011;3:702-14. doi: 10.1093/gbe/evr064. Epub 2011 Jul 6. Genome Biol Evol. 2011. PMID: 21737395 Free PMC article.
-
Evolutionary dynamics of insect symbiont associations.Trends Ecol Evol. 2007 Dec;22(12):625-7. doi: 10.1016/j.tree.2007.08.013. Epub 2007 Nov 5. Trends Ecol Evol. 2007. PMID: 17981362
-
Insect symbionts as hidden players in insect-plant interactions.Trends Ecol Evol. 2012 Dec;27(12):705-11. doi: 10.1016/j.tree.2012.08.013. Epub 2012 Sep 15. Trends Ecol Evol. 2012. PMID: 22985943 Review.
-
Genomics of intracellular symbionts in insects.Int J Med Microbiol. 2010 Jun;300(5):271-8. doi: 10.1016/j.ijmm.2009.12.001. Epub 2010 Jan 25. Int J Med Microbiol. 2010. PMID: 20093081 Review.
Cited by
-
A global characterization and identification of multifunctional enzymes.PLoS One. 2012;7(6):e38979. doi: 10.1371/journal.pone.0038979. Epub 2012 Jun 18. PLoS One. 2012. PMID: 22723914 Free PMC article.
-
Microbial lifestyle and genome signatures.Curr Genomics. 2012 Apr;13(2):153-62. doi: 10.2174/138920212799860698. Curr Genomics. 2012. PMID: 23024607 Free PMC article.
-
Metabolic classification of microbial genomes using functional probes.BMC Genomics. 2012 Apr 27;13:157. doi: 10.1186/1471-2164-13-157. BMC Genomics. 2012. PMID: 22537274 Free PMC article.
-
Computational prediction of genomic functional cores specific to different microbes.J Mol Evol. 2006 Dec;63(6):733-46. doi: 10.1007/s00239-005-0250-9. Epub 2006 Nov 10. J Mol Evol. 2006. PMID: 17103060
-
Unique trajectory of gene family evolution from genomic analysis of nearly all known species in an ancient yeast lineage.bioRxiv [Preprint]. 2024 Jun 6:2024.06.05.597512. doi: 10.1101/2024.06.05.597512. bioRxiv. 2024. PMID: 38895429 Free PMC article. Preprint.
References
-
- Adams, M. D., S. E. Celniker, R. A. Holt, C. A. Evans, J. D. Gocayne, P. G. Amanatides, S. E. Scherer, P. W. Li, R. A. Hoskins, R. F. Galle, R. A. George, S. E. Lewis, S. Richards, M. Ashburner, S. N. Henderson, G. G. Sutton, J. R. Wortman, M. D. Yandell, Q. Zhang, L. X. Chen, R. C. Brandon, Y. H. Rogers, R. G. Blazej, M. Champe, B. D. Pfeiffer, K. H. Wan, C. Doyle, E. G. Baxter, G. Helt, C. R. Nelson, G. L. Gabor, J. F. Abril, A. Agbayani, H. J. An, C. Andrews-Pfannkoch, D. Baldwin, R. M. Ballew, A. Basu, J. Baxendale, L. Bayraktaroglu, E. M. Beasley, K. Y. Beeson, P. V. Benos, B. P. Berman, D. Bhandari, S. Bolshakov, D. Borkova, M. R. Botchan, J. Bouck, P. Brokstein, P. Brottier, K. C. Burtis, D. A. Busam, H. Butler, E. Cadieu, A. Center, I. Chandra, J. M. Cherry, S. Cawley, C. Dahlke, L. B. Davenport, P. Davies, B. de Pablos, A. Delcher, Z. Deng, A. D. Mays, I. Dew, S. M. Dietz, K. Dodson, L. E. Doup, M. Downes, S. Dugan-Rocha, B. C. Dunkov, P. Dunn, K. J. Durbin, C. C. Evangelista, C. Ferraz, S. Ferriera, W. Fleischmann, C. Fosler, A. E. Gabrielian, N. S. Garg, W. M. Gelbart, K. Glasser, A. Glodek, F. Gong, J. H. Gorrell, Z. Gu, P. Guan, M. Harris, N. L. Harris, D. Harvey, T. J. Heiman, J. R. Hernandez, J. Houck, D. Hostin, K. A. Houston, T. J. Howland, M. H. Wei, C. Ibegwam, M. Jalali, F. Kalush, G. H. Karpen, Z. Ke, J. A. Kennison, K. A. Ketchum, B. E. Kimmel, C. D. Kodira, C. Kraft, S. Kravitz, D. Kulp, Z. Lai, P. Lasko, Y. Lei, A. A. Levitsky, J. Li, Z. Li, Y. Liang, X. Lin, X. Liu, B. Mattei, T. C. McIntosh, M. P. McLeod, D. McPherson, G. Merkulov, N. V. Milshina, C. Mobarry, J. Morris, A. Moshrefi, S. M. Mount, M. Moy, B. Murphy, L. Murphy, D. M. Muzny, D. L. Nelson, D. R. Nelson, K. A. Nelson, K. Nixon, D. R. Nusskern, J. M. Pacleb, M. Palazzolo, G. S. Pittman, S. Pan, J. Pollard, V. Puri, M. G. Reese, K. Reinert, K. Remington, R. D. Saunders, F. Scheeler, H. Shen, B. C. Shue, I. Siden-Kiamos, M. Simpson, M. P. Skupski, T. Smith, E. Spier, A. C. Spradling, M. Stapleton, R. Strong, E. Sun, R. Svirskas, C. Tector, R. Turner, E. Venter, A. H. Wang, X. Wang, Z. Y. Wang, D. A. Wassarman, G. M. Weinstock, J. Weissenbach, S. M. Williams, T. Woodage, K. C. Worley, D. Wu, S. Yang, Q. A. Yao, J. Ye, R. F. Yeh, J. S. Zaveri, M. Zhan, G. Zhang, Q. Zhao, L. Zheng, X. H. Zheng, F. N. Zhong, W. Zhong, X. Zhou, S. Zhu, X. Zhu, H. O. Smith, R. A. Gibbs, E. W. Myers, G. M. Rubin and J. C. Venter. 2000. The genome sequence of Drosophila melanogaster. Science 287:2185-2195. - PubMed
-
- Akman, L., A. Yamashita, H. Watanabe, K. Oshima, T. Shiba, M. Hattori, and S. Aksoy. 2002. Genome sequence of the endocellular obligate symbiont of tsetse flies, Wigglesworthia glossinidia. Nat. Genet. 32:402-407. - PubMed
-
- Allikmets, R., B. Gerrard, D. Court, and M. Dean. 1993. Cloning and organization of the abc and mdl genes of Escherichia coli: relationship to eukaryotic multidrug resistance. Gene 136:231-236. - PubMed
-
- Andersson, S. G. E., A. Zomorodipour, J. O. Andersson, T. Sicheritz-Ponten, U. C. M. Alsmark, R. M. Podowski, A. K. Naslund, A. S. Eriksson, H. H. Winkler, and C. G. Kurland. 1998. The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature 396:133-140. - PubMed
-
- Barancin, C. E., J. C. Smoot, R. H. Findlkay, and L. A. Actis. 1998. Plasmid- mediated histamine biosynthesis in the bacterial fish pathogen Vibrio anguillarum. Plasmid 39:235-244. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
