

Review
doi: 10.1128/MCB.24.24.10505-10514.2004.
Influence of RNA secondary structure on the pre-mRNA splicing process
Affiliations
- PMID: 15572659
- PMCID: PMC533984
- DOI: 10.1128/MCB.24.24.10505-10514.2004
Item in Clipboard
Review
Influence of RNA secondary structure on the pre-mRNA splicing process
Mol Cell Biol.
2004 Dec.
No abstract available
Figures
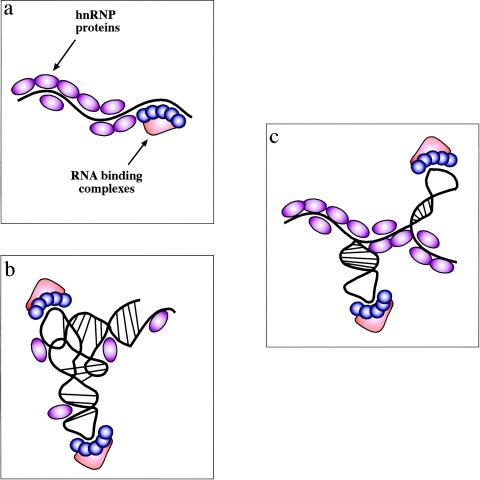
Experimental models of RNA secondary structures in mRNAs. There are three possible experimental models of RNA secondary structures in mRNAs. (a) The first one is represented by the view that hnRNP proteins bind the mRNA as it gets transcribed by RNA polymerase II and keep it in a largely linear conformation. In this case, binding of specific factors is regulated only by the competitive advantage provided by sequence-specific interactions over the generic RNA binding affinities of all hnRNP proteins. (b) The opposite situation is one where the drive to form RNA secondary and tertiary structures is stronger than the ability of RNA-binding proteins to prevent it (and maybe even stabilized by these proteins). In this case, the role played by generic RNA-binding proteins is severely reduced and specific complexes can bind through a mix of sequence-specific and structure-specific recognition. (c) Between these two models is a situation that should encompass many cellular mRNAs. In this case, the potential “ironing” of the mRNA by its weak or aspecific interactions with hnRNPs can indeed maintain the mRNA in a largely linear conformation. However, in particular regions the mRNA is still able to form localized RNA structures which might represent, together with the nucleotide sequence, preferential binding sites for specific nuclear complexes. Of course, given the enormous variety of mRNAs produced by the cell these models cannot be considered mutually exclusive, although there is probably a distinct preference for the model in panel c.

Effects of RNA secondary structure on protein binding motifs. (a) Loosely conserved RNA binding sequences can be induced to display a uniform binding surface following the formation of a hairpin structure. The example chosen is the protein HuR, which binds to AU- and U-rich mRNA regions and affects their stability and translation. Identification of a large set of target sequences has shown that the they consist of two complementary 6-bp motifs (highlighted in red and violet) and a loop region which contains only a conserved U residue (highlighted in blue) (68). (b) Splicing factors are also capable of affecting RNA spatial distribution (besides being affected by it). For example, binding of SF-1, U2AF65, and U2AF35 to a small 62-nt RNA containing a branch point sequence (BPS), polypyrimidine tract (PPT), and 3′ splice site (3'ss) has been shown by hydroxyl radical iron-EDTA probing to bend the RNA in such a way as to bring together the 3′ splice site and branch point region, thus helping the formation of the splicing commitment complex (59).
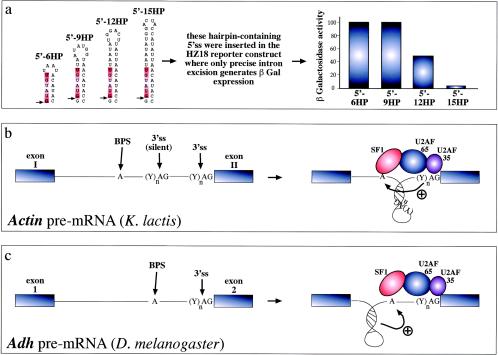
RNA structural elements and splicing efficiency. (a) Series of short artificial hairpins (HP) of increasing length containing a 5′ splice site (indicated by an arrow, while the consensus sequence is highlighted in red). These constructs were engineered in the yeast RP51A intron and assayed for functionality in a reporter construct which contains a wild-type intron inserted in the lacZ gene. In this construct, only precise excision of the intron generates β-galactosidase expression. Following transfection in yeast cells, it was determined that longer hairpins have an increasing ability to sequester the donor site and inhibit the early steps of spliceosome assembly (43). Hairpins can also change the relative distances of splicing regulatory elements and thus affect the final outcome. For example, in the yeast actin intron (b) the branch-point sequence (BPS) is located far away from the 3′splice site (3'ss), which is also preceded by a silent 3'ss (silent) that is not normally used by the splicing machinery. The reason for this lies in the proposed folding of the region between the BPS and the correct 3'ss in a hairpin structure. The function of this structure would be twofold: to bring the BPS into working range of the correct 3'ss and to sequester the silent 3'ss, preventing its use by the splicing machinery (33). Finally, hairpin structure formation near the branch point of the Adh gene intron 1 (3c) has been recently proposed to play an active role in splicing through a distinct mechanism. In this case, hairpin formation has been proposed to force the branch point sequence (BPS) into an unpaired conformation that would be better recognized by the splicing machinery (23).
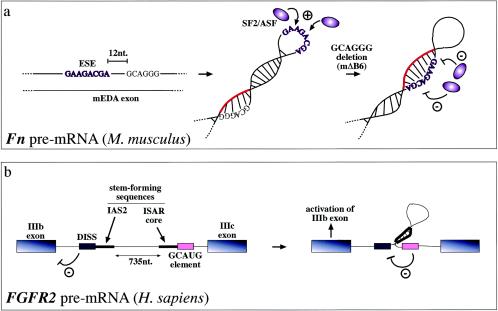
Steric hindrance has also been found to occur in ESE sequences, which promote recognition of the correct 3′ splice sites and 5′ splice sites. In the case of the mouse (and human) fibronectin EDA exons, secondary structural elements can stabilize the conformation of the ESE sequence and enhance its SR protein binding capabilities (13, 77). For example, mutations that do not directly affect the mouse ESE sequence have been demonstrated to cause a conformational change in this region (from a loop to a stem) which hinders SF2/ASF protein binding and in this way abolishes exon recognition (a). Highlighted in red is the RNA region which, following the mΔB6 deletion, blocks the ESE sequence. Alternatively, RNA secondary structures can also function on ESE/ESS regulatory regions indirectly. For example, in the FGFR2 gene they contribute to regulating the inclusion of the mutually exclusive IIIb (expressed in epithelial cells) and IIIc (expressed in mesenchymal cells) exons. In this case, the function of the stem structure formed by the intronic activating sequence 2 (IAS2) and intronic splicing activator and repressor (ISAR) element would be that of approximating an inhibitory intronic sequence (a GCAUG-rich sequence) relative to the distal intronic splicing silencer (DISS) element, which would normally repress exon IIIb inclusion (4). Inactivation of this element would then lead to activation of exon IIIb splicing in epithelial cells (b).
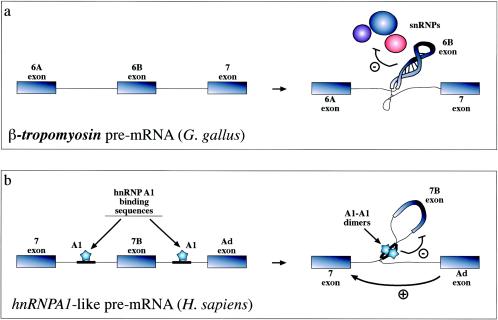
Recognition of exons can also be inhibited by formation of extensive RNA secondary structures such as the one shown (a) for the chicken β-tropomyosin gene. In this case, the 6B exonic sequence and its surrounding intronic sequences fold upon themselves in a complex structure that has the ability to affect the interaction of all snRNPs (U1 to U6) with the pre-mRNA (91). Formation of RNA structures that can “loop out” an entire pre-mRNA has also been described to occur in the hnRNPA1 pre-mRNA. However, in this case inhibition is mediated by proteins (hnRNP A1 itself) binding on either side of the exon and interacting with each other (b) (79). The way these structures have been proposed to act is through steric hindrance of the looped-out splice sites (which even if it does not hinder U1snRNP binding may be incompatible with later splicing events). Alternatively, or in addition, these structures might function by simply providing a competitive advantage for distal 5′ splice sites which are moved closer to the acceptor sequence.
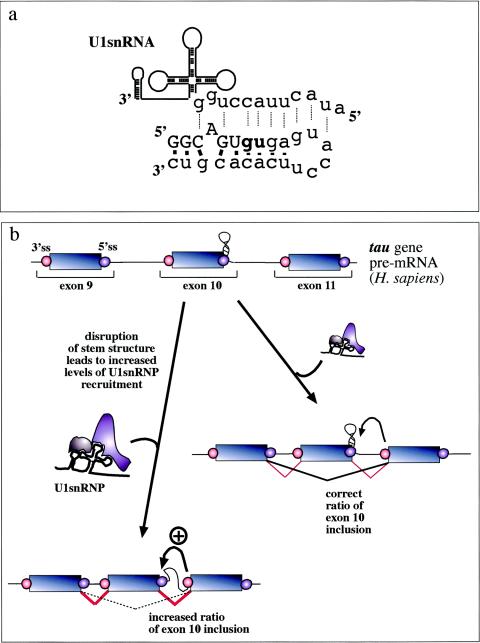
Involvement of RNA structural elements in human disease. In the alternatively spliced tau exon 10, a stem-loop element regulates the accessibility of the 5′ splice site by U1snRNP. (a) What NMR studies have uncovered (100). This element consists of a stem-loop structure involving the 5′ exon/intron junction (uppercase/lowercase) from −5 to +19 which is capped by a flexible loop of 6 bp. Base pairings between the nucleotides in this region (solid bars) prevent recognition by U1snRNA (dotted lines). (b) Mutations which disrupt the structural element result in an increased accessibility of this region and an increased binding of U1snRNP to the donor site. As a consequence, exon recognition is improved and the resulting alterations in the balance between exon 10+ and exon 10− transcripts inside the cell have been proposed to lead to frontotemporal dementia and parkinsonism linked to chromosome 17 (46, 53, 100, 101, 108).
Similar articles
-
Secondary structure of the yeast Saccharomyces cerevisiae pre-U3A snoRNA and its implication for splicing efficiency.RNA. 1996 Nov;2(11):1079-93. RNA. 1996. PMID: 8903339 Free PMC article.
-
Genetics. Minor splicing, disrupted.Science. 2011 Apr 8;332(6026):184-5. doi: 10.1126/science.1205503. Science. 2011. PMID: 21474744 No abstract available.
-
U2 toggles iteratively between the stem IIa and stem IIc conformations to promote pre-mRNA splicing.Genes Dev. 2007 Apr 1;21(7):821-34. doi: 10.1101/gad.1536107. Genes Dev. 2007. PMID: 17403782 Free PMC article.
-
RNA Splicing by the Spliceosome.Annu Rev Biochem. 2020 Jun 20;89:359-388. doi: 10.1146/annurev-biochem-091719-064225. Epub 2019 Dec 3. Annu Rev Biochem. 2020. PMID: 31794245 Review.
-
How Is Precursor Messenger RNA Spliced by the Spliceosome?Annu Rev Biochem. 2020 Jun 20;89:333-358. doi: 10.1146/annurev-biochem-013118-111024. Epub 2019 Dec 9. Annu Rev Biochem. 2020. PMID: 31815536 Review.
Cited by
-
Positional mapping and candidate gene analysis of the mouse Ccs3 locus that regulates differential susceptibility to carcinogen-induced colorectal cancer.PLoS One. 2013;8(3):e58733. doi: 10.1371/journal.pone.0058733. Epub 2013 Mar 14. PLoS One. 2013. PMID: 23516545 Free PMC article.
-
Splicing regulation in spinal muscular atrophy by an RNA structure formed by long-distance interactions.Ann N Y Acad Sci. 2015 Apr;1341:176-87. doi: 10.1111/nyas.12727. Epub 2015 Feb 27. Ann N Y Acad Sci. 2015. PMID: 25727246 Free PMC article.
-
Conserved long-range base pairings are associated with pre-mRNA processing of human genes.Nat Commun. 2021 Apr 16;12(1):2300. doi: 10.1038/s41467-021-22549-7. Nat Commun. 2021. PMID: 33863890 Free PMC article.
-
EDA Fibronectin in Keloids Create a Vicious Cycle of Fibrotic Tumor Formation.J Invest Dermatol. 2015 Jul;135(7):1714-1718. doi: 10.1038/jid.2015.155. J Invest Dermatol. 2015. PMID: 26066891
-
Pyrvinium pamoate changes alternative splicing of the serotonin receptor 2C by influencing its RNA structure.Nucleic Acids Res. 2013 Apr 1;41(6):3819-32. doi: 10.1093/nar/gkt063. Epub 2013 Feb 7. Nucleic Acids Res. 2013. PMID: 23393189 Free PMC article.
References
-
- Adams, M. D., D. Z. Rudner, and D. C. Rio. 1996. Biochemistry and regulation of pre-mRNA splicing. Curr. Opin. Cell Biol. 8:331-339. - PubMed
-
- Antson, A. A. 2000. Single-stranded-RNA binding proteins. Curr. Opin. Struct. Biol. 10:87-94. - PubMed
-
- Balvay, L., D. Libri, and M. Y. Fiszman. 1993. Pre-mRNA secondary structure and the regulation of splicing. Bioessays 15:165-169. - PubMed
-
- Black, D. L. 1991. Does steric interference between splice sites block the splicing of a short c-src neuron-specific exon in non-neuronal cells? Genes Dev. 5:389-402. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources