

doi: 10.1101/gad.303304.
Slik Sterile-20 kinase regulates Moesin activity to promote epithelial integrity during tissue growth
Affiliations
- PMID: 15371338
- PMCID: PMC517517
- DOI: 10.1101/gad.303304
Item in Clipboard
Slik Sterile-20 kinase regulates Moesin activity to promote epithelial integrity during tissue growth
Genes Dev.
.
Abstract
The Drosophila Sterile-20 kinase Slik promotes tissue growth during development by stimulating cell proliferation and by preventing apoptosis. Proliferation within an epithelial sheet requires dynamic control of cellular architecture. Epithelial integrity fails in slik mutant imaginal discs. Cells leave the epithelium and undergo apoptosis. The abnormal behavior of slik mutant cells is due to failure to phosphorylate and activate Moesin, which leads to excess Rho1 activity. This is distinct from Slik's effects on cell proliferation, which are mediated by Raf. Thus Slik acts via distinct pathways to coordinate cell proliferation with epithelial cell behavior during tissue growth.
Figures
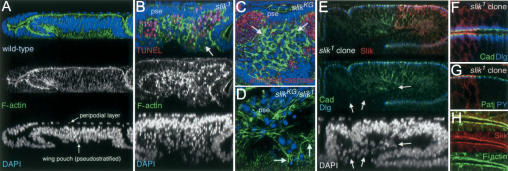
Epithelial integrity defects in Slik mutants. (A) Optical cross-section of a wild-type wing imaginal disc showing the presumptive wing, labeled to visualize F-actin (green) and nuclei (DAPI, blue). Note the multilayered nuclei in the pseudostratified wing pouch area, and the transition from columnar to squamous epithelial organization in the overlying peripodial layer. F-actin is enriched at the apical junctional complexes. (B) Optical cross-section of a slik1 homozygous null mutant wing disc. Apoptotic cells were labeled by TUNEL (red). The pseudostratified epithelium (pse) is abnormally thin. The space beneath the basal surface of the disc is full of extruded cells; many are TUNEL-positive with pyknotic nuclei. Clusters of TUNEL-negative live cells are also extruded (arrow). Note also a few apoptotic cells in the epithelial layer. (C,D) Horizontal optical sections taken just below the wing pouch epithelium of slikKG4837 hypomorphic mutant and slikKG4837/slik1 mutant discs. Apoptotic cells are labeled with antibody to activated caspase 3 (red in C). slik mutant cells with abnormally high levels of F-actin (arrows in C) lose their epithelial character and are extruded from the basal surface of the epithelium (pse, viewed in cross-section). Arrows in D indicate F-actin-rich filopodia-like extensions in a cluster of cells migrating out of the epithelium (pse). (E-G) Cross-sections through wing disc epithelia with large clones of slik1 mutant cells. Slik protein (red) is absent from the clones. (E,F) E-Cadherin (green) and Discs large protein (blue) are localized normally in mutant cells in the epithelium, but are not localized in extruded cells (arrows in E). (F) Slik is apical to E-Cadherin and Discs large. (G) Slik is apical to the Patj protein (green) and to adherens junctions labeled by concentrated Phosphotyrosine (blue). (H) Slik and F-actin colocalize apically in wild-type cells in an optical section that crosses a fold in the disc.
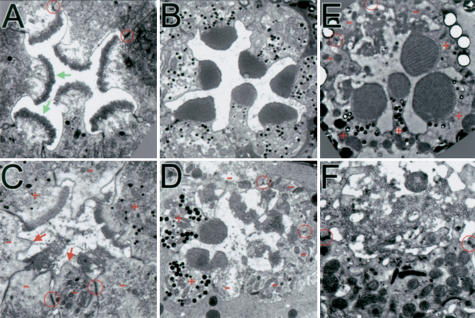
Defects in rhabdomere differentiation in slik mutant photoreceptors. (A,B) Transmission electron microscopy (TEM) sections of single wild-type ommatidia. (A) 70% PD. Some adherens junctions are indicated (red circles). Green arrows indicate forming microvilli arrays. (B) 95% PD. (C-F) Ommatidia with slik1 mutant clones. Wild-type cells have pigment granules (+). (-) Mutant cells; (red arrows) apical membranes lacking microvilli. (C) 70% PD. (D) 95% PD. (E,F) 18-day-old adult. (F) Detail of the disorganized microvilli.
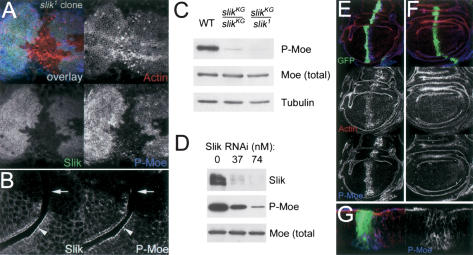
Slik kinase is required for regulatory phosphorylation of Moesin. (A) Projection of a series of optical sections through the apical region of a wing disc with a large slik1 mutant clone. Thr 556-phosphorylated Moesin (P-Moe) was detected with a phospho-specific antibody (Polesello et al. 2002). P-Moe (blue) is reduced in cells lacking Slik protein (green). F-actin is shown in red. (B) An optical section through another clone crossing a fold shows two cell layers with their apical surfaces apposed. Slik and P-Moesin colocalize apically in wild-type cells (arrowheads), but are absent in slik mutant cells (arrows). (C) Immunoblot of extracts from wild-type (WT), homozygous slikKG4837, and slikKG4837/slik1 mutant wing discs. (D) Immunoblot of extracts from S2 cells treated with increasing concentrations of double-stranded RNA for Slik. (E-G) Wing discs expressing GFP (green) and wild-type (E,G) or catalytically inactive (F) forms of the Slik kinase domain under ptcGal4 control and labeled for F-actin (red) and PMoesin (blue). (G) Cross-section of the disc in E. The Slik kinase domain is not apically localized and causes relocalization of P-Moesin from the apical surface.
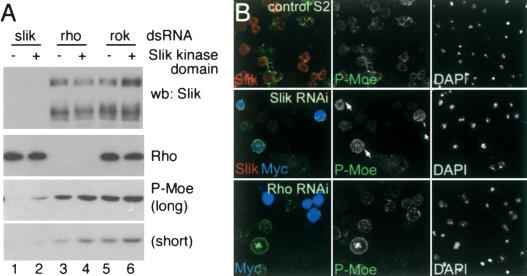
Role of Rho and Slik in Moesin phosphorylation. (A) Immunoblots of lysates from S2 cells treated with double-stranded RNA for Slik, Rho, or Rok. (+) Cells transfected to express the Slik kinase domain. (Upper panel) Slik protein was depleted by RNAi. The transfected kinase domain is not detected by the Slik antibody. (Second panel) Rho protein was depleted by RNAi. (Lower two panels) Long and short exposures of immunoblots showing levels of P-Moesin. (B) S2 cells labeled with anti-Slik (red), anti-P-Moe (green), and anti-Myc to visualize the transfected Myc-tagged Slik kinase domain (blue). Nuclei labeled with DAPI are shown at right. (Upper panels) Control S2 cells. (Middle panels) Cells treated with Slik dsRNA. Slik and P-Moe levels were strongly reduced. Few untransfected cells had high levels of P-Moe, but cells expressing the Slik kinase domain did (arrows). (Lower panels) Cells treated with Rho dsRNA. P-Moe levels were comparable to wild-type cells.
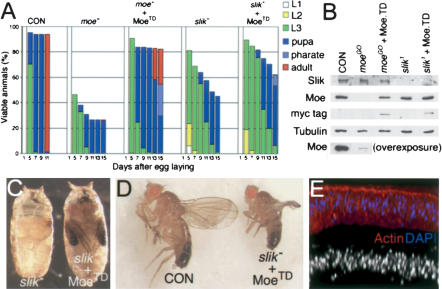
Partial rescue of slik mutants by activated Moesin. (A) Histograms showing the proportion of animals at different larval, pupal, and adult stages as a function of age. (moe-) moeGO323/Y mutant; (MoeTD) Myc-epitope tagged Thr 556 Asp phosphomimetic form of Moesin; (slik-) slik1/slik1. Numbers of larvae: CON, n = 80; moe-, n = 97; moe- + MoeTD, n = 191; slik-, n = 80; slik- + MoeTD, n = 280. (B) Immunoblots of imaginal disc lysates as in A. (C) slik1 mutant pupa and slik1 armGAL4 UAS-MoeTD mutant rescued to pharate adult stage. Eyes (orange), wings (black), and legs are seen. (D) A rare adult survivor (1/280) of the rescued genotype. (E) Optical cross-section of a slik1 armGAL4 UAS-MoeTD wing disc.
Similar articles
-
The Drosophila sterile-20 kinase slik controls cell proliferation and apoptosis during imaginal disc development.PLoS Biol. 2003 Nov;1(2):E35. doi: 10.1371/journal.pbio.0000035. Epub 2003 Nov 17. PLoS Biol. 2003. PMID: 14624240 Free PMC article.
-
Sip1, the Drosophila orthologue of EBP50/NHERF1, functions with the sterile 20 family kinase Slik to regulate Moesin activity.J Cell Sci. 2010 Apr 1;123(Pt 7):1099-107. doi: 10.1242/jcs.059469. Epub 2010 Mar 9. J Cell Sci. 2010. PMID: 20215404 Free PMC article.
-
Extrusion of cells with inappropriate Dpp signaling from Drosophila wing disc epithelia.Science. 2005 Mar 18;307(5716):1789-90. doi: 10.1126/science.1104784. Science. 2005. PMID: 15774763
-
Filling out the Hippo pathway.Nat Rev Mol Cell Biol. 2007 Aug;8(8):613-21. doi: 10.1038/nrm2221. Nat Rev Mol Cell Biol. 2007. PMID: 17622252 Review.
-
Cytokinesis genetics takes wing.Trends Genet. 1990 Oct;6(10):309-10. Trends Genet. 1990. PMID: 2281526 Review. No abstract available.
Cited by
-
Effector-mediated ERM activation locally inhibits RhoA activity to shape the apical cell domain.J Cell Biol. 2021 Jun 7;220(6):e202007146. doi: 10.1083/jcb.202007146. J Cell Biol. 2021. PMID: 33836044 Free PMC article.
-
Molecular networks linked by Moesin drive remodeling of the cell cortex during mitosis.J Cell Biol. 2011 Oct 3;195(1):99-112. doi: 10.1083/jcb.201106048. J Cell Biol. 2011. PMID: 21969469 Free PMC article.
-
Slik and the receptor tyrosine kinase Breathless mediate localized activation of Moesin in terminal tracheal cells.PLoS One. 2014 Jul 25;9(7):e103323. doi: 10.1371/journal.pone.0103323. eCollection 2014. PLoS One. 2014. PMID: 25061859 Free PMC article.
-
Isoform-specific functions of Mud/NuMA mediate binucleation of Drosophila male accessory gland cells.BMC Dev Biol. 2014 Dec 20;14:46. doi: 10.1186/s12861-014-0046-5. BMC Dev Biol. 2014. PMID: 25527079 Free PMC article.
-
Shot and Patronin polarise microtubules to direct membrane traffic and biogenesis of microvilli in epithelia.J Cell Sci. 2016 Jul 1;129(13):2651-9. doi: 10.1242/jcs.189076. Epub 2016 May 26. J Cell Sci. 2016. PMID: 27231092 Free PMC article.
References
-
- Bilder D., Li, M., and Perrimon, N. 2000. Cooperative regulation of cell polarity and growth by Drosophila tumor suppressors. Science 289: 113-116. - PubMed
-
- Dan I., Watanabe, N.M., and Kusumi, A. 2001. The Ste20 group kinases as regulators of MAP kinase cascades. Trends Cell Biol. 11: 220-230. - PubMed
-
- Denker S.P., Huang, D.C., Orlowski, J., Furthmayr, H., and Barber, D.L. 2000. Direct binding of the Na-H exchanger NHE1 to ERM proteins regulates the cortical cytoskeleton and cell shape independently of H(+) translocation. Mol. Cell 6: 1425-1436. - PubMed
-
- Doi Y., Itoh, M., Yonemura, S., Ishihara, S., Takano, H., Noda, T., and Tsukita, S. 1999. Normal development of mice and unimpaired cell adhesion/cell motility/actin-based cytoskeleton without compensatory up-regulation of ezrin or radixin in moesin gene knockout. J. Biol. Chem. 274: 2315-2321. - PubMed
-
- Evan G. and Littlewood, T. 1998. A matter of life and cell death. Science 281: 1317-1322. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous