

The phosphatidylserine receptor has essential functions during embryogenesis but not in apoptotic cell removal
- PMID: 15345036
- PMCID: PMC549712
- DOI: 10.1186/jbiol10
The phosphatidylserine receptor has essential functions during embryogenesis but not in apoptotic cell removal
Abstract
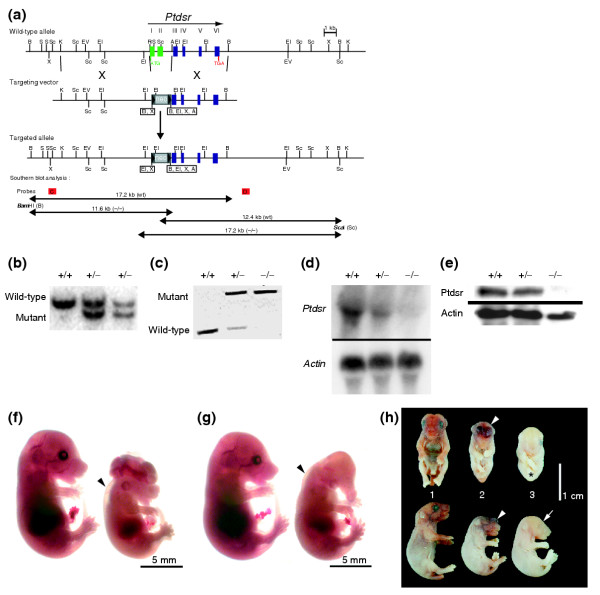
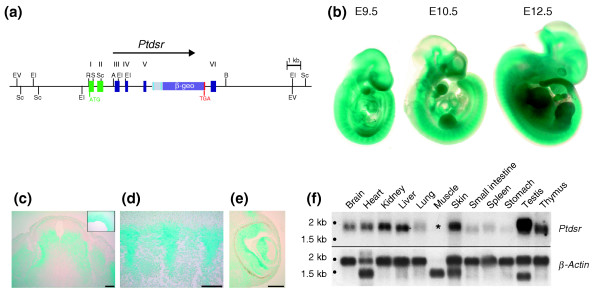
Background: Phagocytosis of apoptotic cells is fundamental to animal development, immune function and cellular homeostasis. The phosphatidylserine receptor (Ptdsr) on phagocytes has been implicated in the recognition and engulfment of apoptotic cells and in anti-inflammatory signaling. To determine the biological function of the phosphatidylserine receptor in vivo, we inactivated the Ptdsr gene in the mouse.
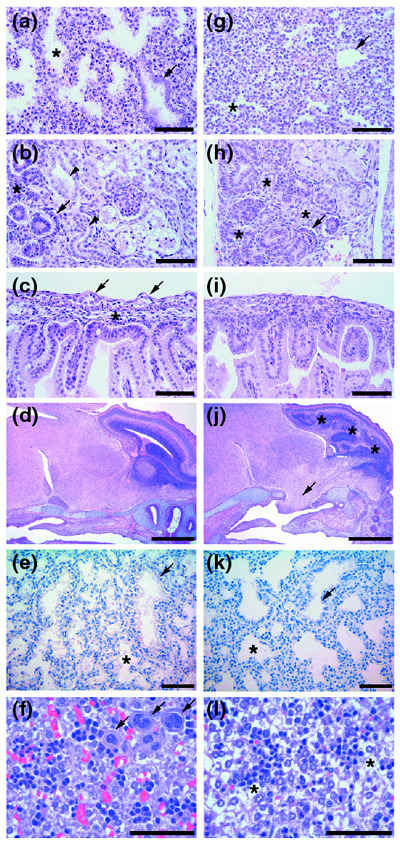
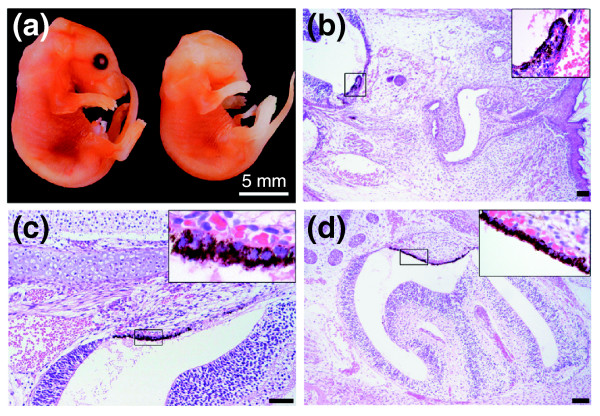
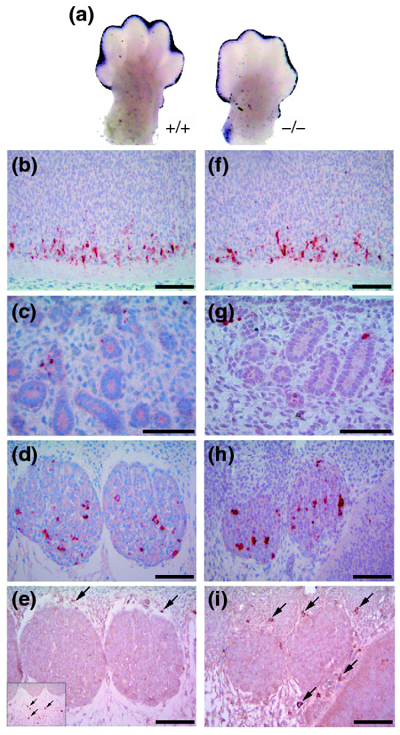
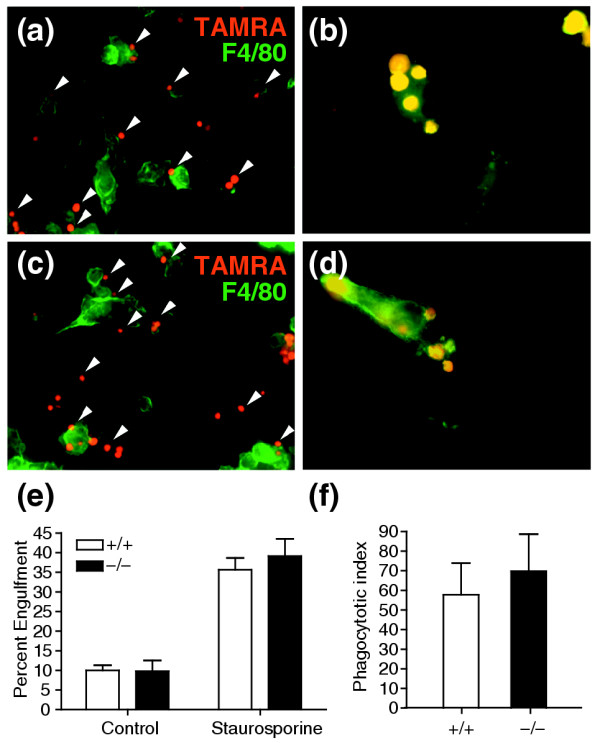
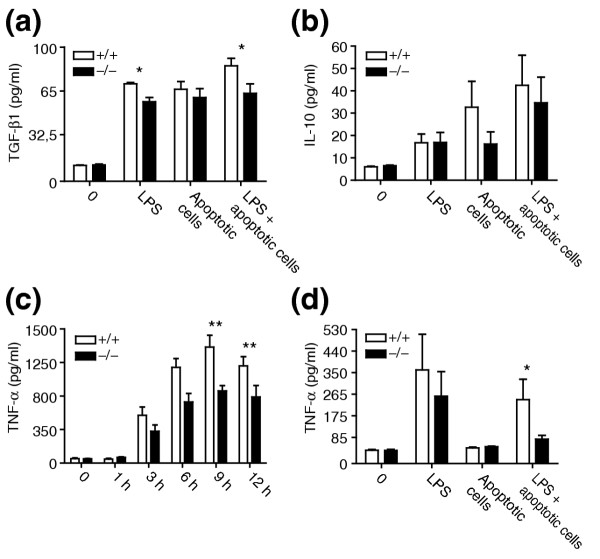
Results: Ablation of Ptdsr function in mice causes perinatal lethality, growth retardation and a delay in terminal differentiation of the kidney, intestine, liver and lungs during embryogenesis. Moreover, eye development can be severely disturbed, ranging from defects in retinal differentiation to complete unilateral or bilateral absence of eyes. Ptdsr -/- mice with anophthalmia develop novel lesions, with induction of ectopic retinal-pigmented epithelium in nasal cavities. A comprehensive investigation of apoptotic cell clearance in vivo and in vitro demonstrated that engulfment of apoptotic cells was normal in Ptdsr knockout mice, but Ptdsr-deficient macrophages were impaired in pro- and anti-inflammatory cytokine signaling after stimulation with apoptotic cells or with lipopolysaccharide.
Conclusion: Ptdsr is essential for the development and differentiation of multiple organs during embryogenesis but not for apoptotic cell removal. Ptdsr may thus have a novel, unexpected developmental function as an important differentiation-promoting gene. Moreover, Ptdsr is not required for apoptotic cell clearance by macrophages but seems to be necessary for the regulation of macrophage cytokine responses. These results clearly contradict the current view that the phosphatidylserine receptor primarily functions in apoptotic cell clearance.
Figures

Similar articles
-
A receptor for phosphatidylserine-specific clearance of apoptotic cells.Nature. 2000 May 4;405(6782):85-90. doi: 10.1038/35011084. Nature. 2000. PMID: 10811223
-
Clearance of apoptotic cells is not impaired in mouse embryos deficient in class A scavenger receptor types I and II (CD204).Dev Dyn. 2005 Jan;232(1):67-74. doi: 10.1002/dvdy.20206. Dev Dyn. 2005. PMID: 15580571
-
Phosphatidylserine receptor is required for clearance of apoptotic cells.Science. 2003 Nov 28;302(5650):1560-3. doi: 10.1126/science.1087621. Science. 2003. PMID: 14645847
-
If phosphatidylserine is the death knell, a new phosphatidylserine-specific receptor is the bellringer.Cell Death Differ. 2001 Jun;8(6):582-7. doi: 10.1038/sj.cdd.4400856. Cell Death Differ. 2001. PMID: 11536008 Review.
-
[Research on Molecular Mechanisms of Engulfment of Apoptotic Cells].Yakugaku Zasshi. 2015;135(8):949-54. doi: 10.1248/yakushi.15-00110. Yakugaku Zasshi. 2015. PMID: 26234352 Review. Japanese.
Cited by
-
Protein Hydroxylation Catalyzed by 2-Oxoglutarate-dependent Oxygenases.J Biol Chem. 2015 Aug 21;290(34):20712-20722. doi: 10.1074/jbc.R115.662627. Epub 2015 Jul 7. J Biol Chem. 2015. PMID: 26152730 Free PMC article. Review.
-
Robustness analysis on interspecies interaction network for iron and glucose competition between Candida albicans and zebrafish during infection.BMC Syst Biol. 2014;8 Suppl 5(Suppl 5):S6. doi: 10.1186/1752-0509-8-S5-S6. Epub 2014 Dec 12. BMC Syst Biol. 2014. PMID: 25603810 Free PMC article.
-
Microvascular endothelial cells express a phosphatidylserine receptor: a functionally active receptor for phosphatidylserine-positive erythrocytes.Blood. 2008 Jan 15;111(2):905-14. doi: 10.1182/blood-2007-07-099465. Epub 2007 Oct 2. Blood. 2008. PMID: 17911385 Free PMC article.
-
Phospholipids: key players in apoptosis and immune regulation.Molecules. 2009 Nov 30;14(12):4892-914. doi: 10.3390/molecules14124892. Molecules. 2009. PMID: 20032867 Free PMC article. Review.
-
Erythropoietin requires NF-kappaB and its nuclear translocation to prevent early and late apoptotic neuronal injury during beta-amyloid toxicity.Curr Neurovasc Res. 2005 Dec;2(5):387-99. doi: 10.2174/156720205774962683. Curr Neurovasc Res. 2005. PMID: 16375720 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
