

Junctate is a key element in calcium entry induced by activation of InsP3 receptors and/or calcium store depletion
- PMID: 15302852
- PMCID: PMC1868564
- DOI: 10.1083/jcb.200404079
Junctate is a key element in calcium entry induced by activation of InsP3 receptors and/or calcium store depletion
Abstract
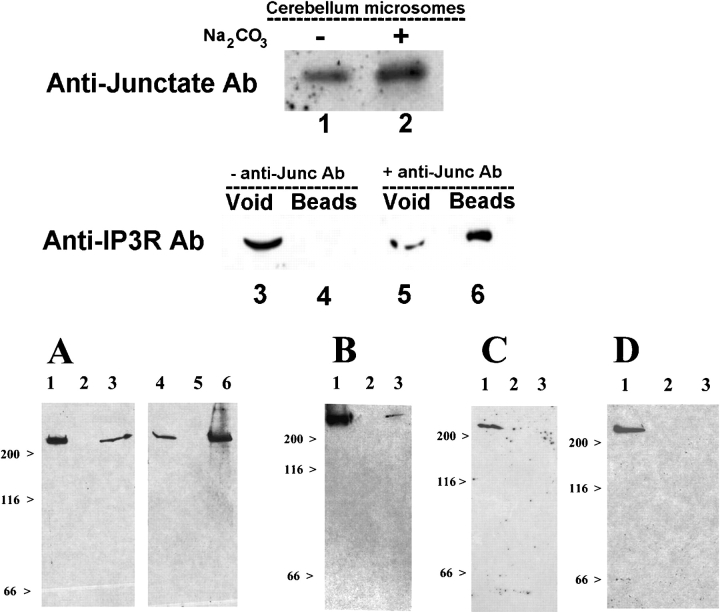
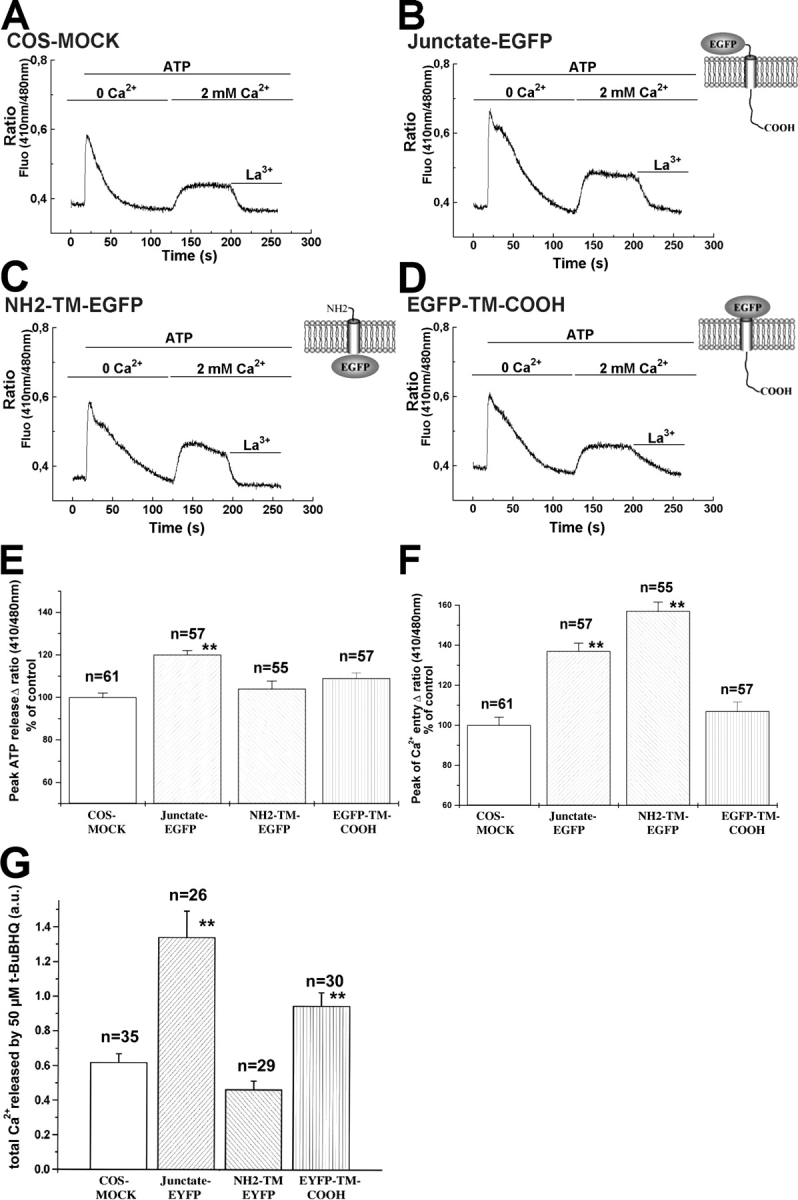
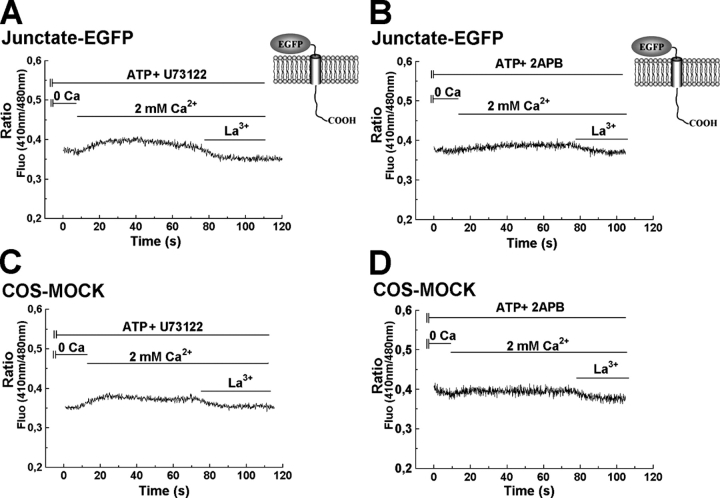
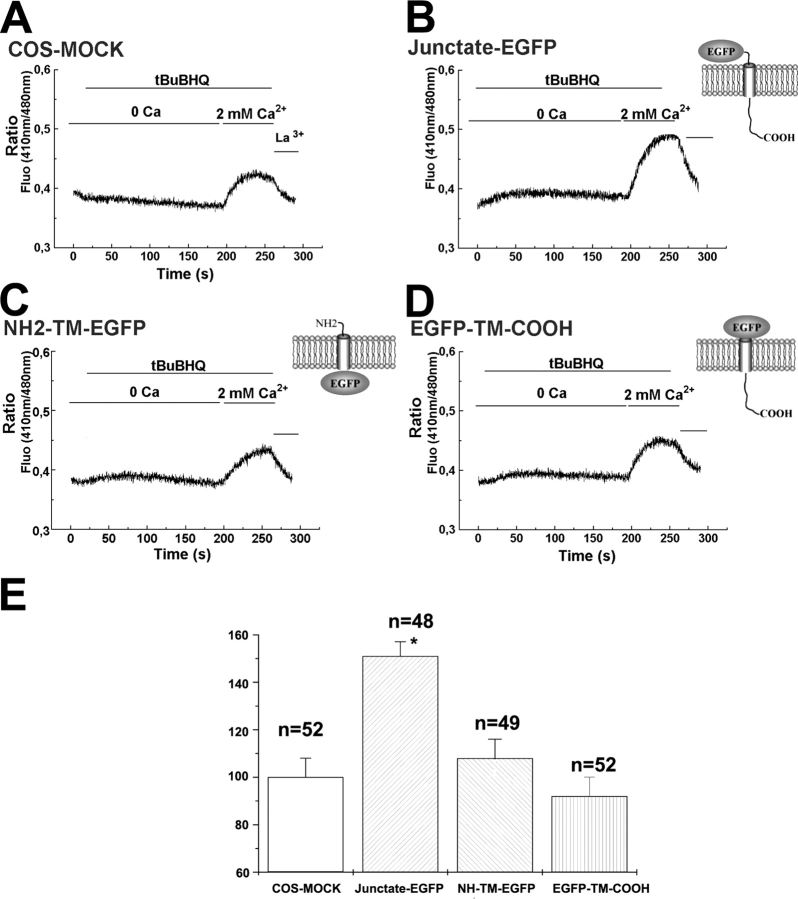
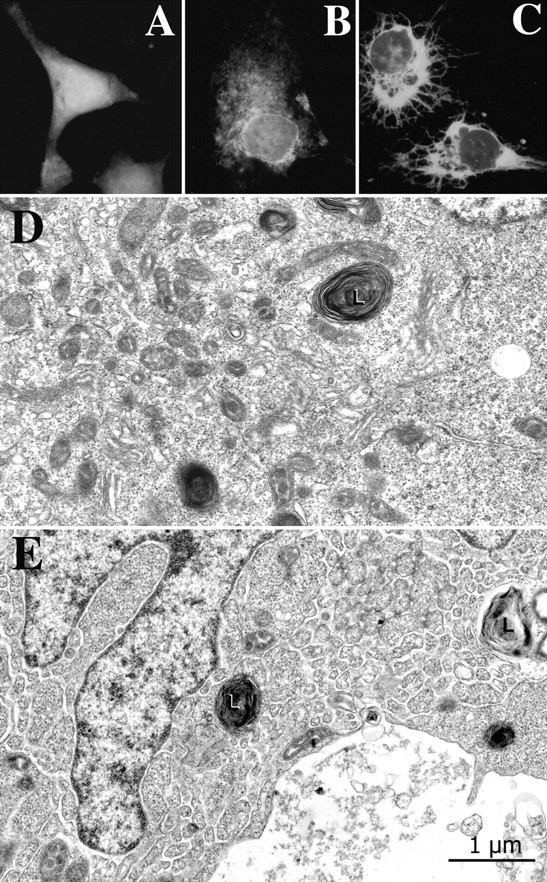
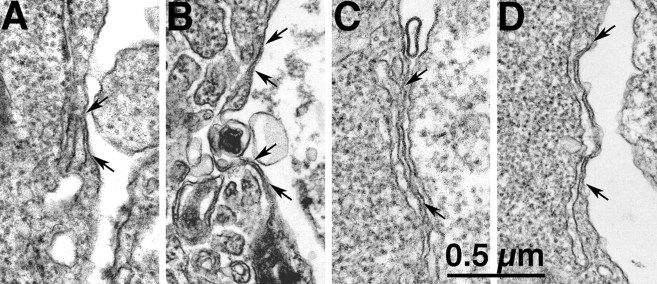
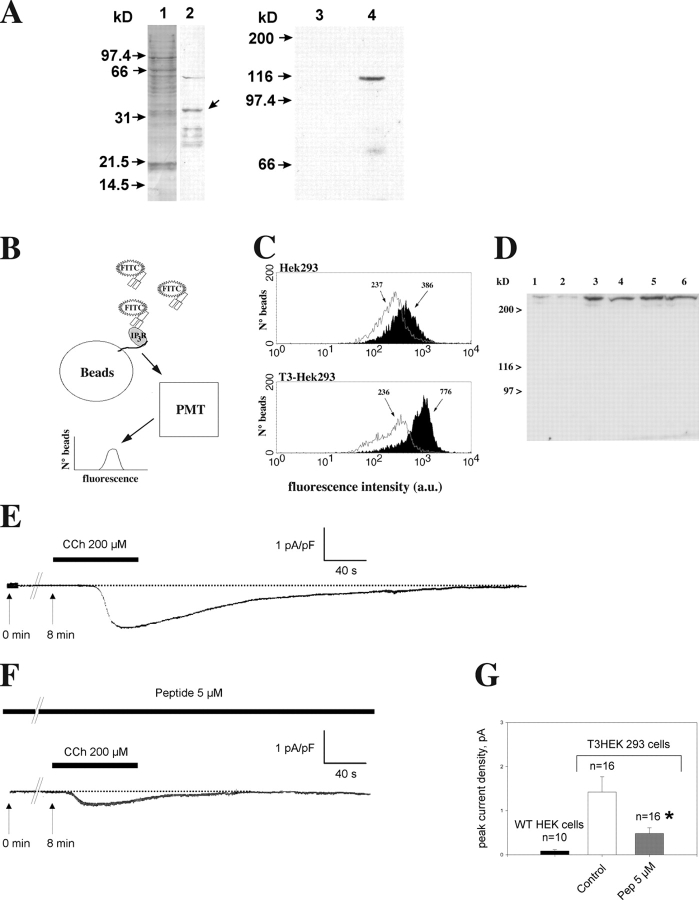
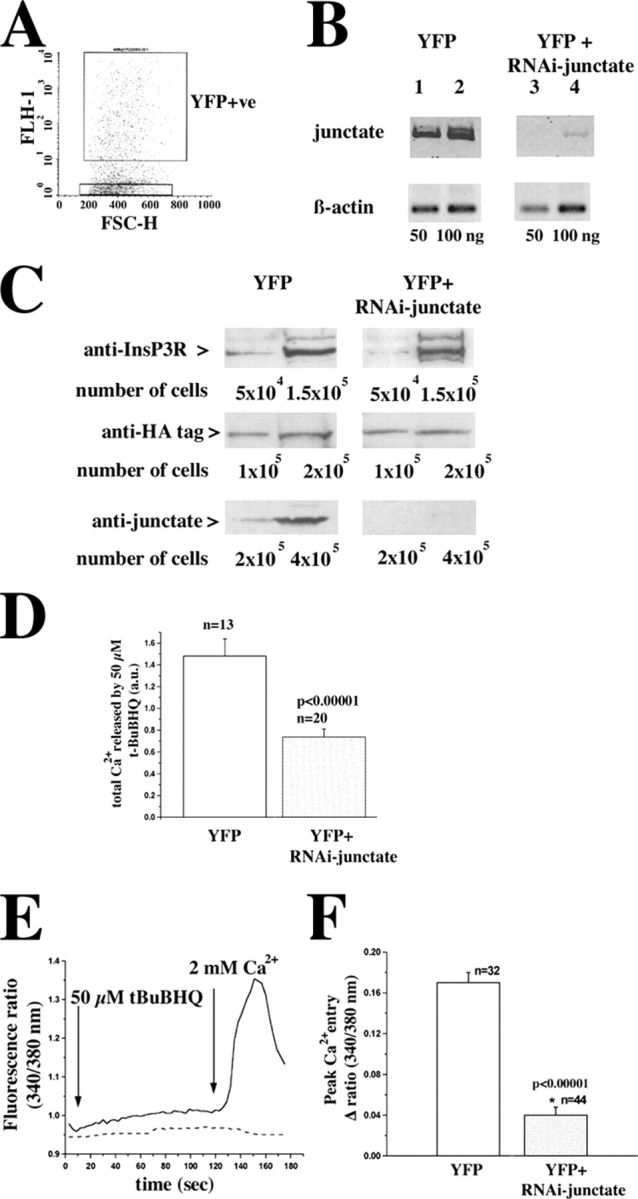
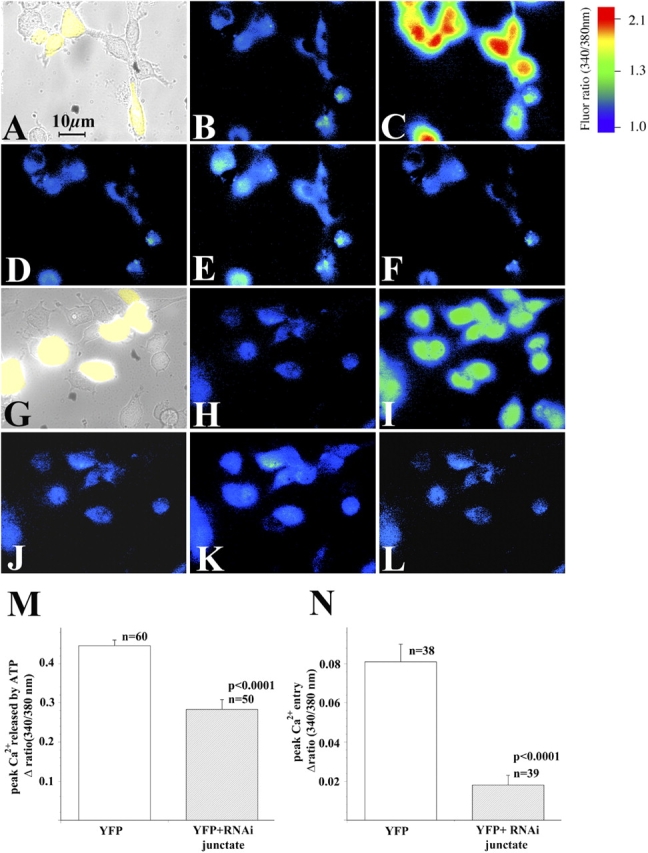
In many cell types agonist-receptor activation leads to a rapid and transient release of Ca(2+) from intracellular stores via activation of inositol 1,4,5 trisphosphate (InsP(3)) receptors (InsP(3)Rs). Stimulated cells activate store- or receptor-operated calcium channels localized in the plasma membrane, allowing entry of extracellular calcium into the cytoplasm, and thus replenishment of intracellular calcium stores. Calcium entry must be finely regulated in order to prevent an excessive intracellular calcium increase. Junctate, an integral calcium binding protein of endo(sarco)plasmic reticulum membrane, (a) induces and/or stabilizes peripheral couplings between the ER and the plasma membrane, and (b) forms a supramolecular complex with the InsP(3)R and the canonical transient receptor potential protein (TRPC) 3 calcium entry channel. The full-length protein modulates both agonist-induced and store depletion-induced calcium entry, whereas its NH(2) terminus affects receptor-activated calcium entry. RNA interference to deplete cells of endogenous junctate, knocked down both agonist-activated calcium release from intracellular stores and calcium entry via TRPC3. These results demonstrate that junctate is a new protein involved in calcium homeostasis in eukaryotic cells.
Figures
Similar articles
-
Molecular cloning, expression, functional characterization, chromosomal localization, and gene structure of junctate, a novel integral calcium binding protein of sarco(endo)plasmic reticulum membrane.J Biol Chem. 2000 Dec 15;275(50):39555-68. doi: 10.1074/jbc.M005473200. J Biol Chem. 2000. PMID: 11007777
-
Junctate, an inositol 1,4,5-triphosphate receptor associated protein, is present in rodent sperm and binds TRPC2 and TRPC5 but not TRPC1 channels.Dev Biol. 2005 Oct 1;286(1):326-37. doi: 10.1016/j.ydbio.2005.08.006. Dev Biol. 2005. PMID: 16153633
-
Agonist-activated Ca2+ influx occurs at stable plasma membrane and endoplasmic reticulum junctions.J Cell Sci. 2010 Dec 1;123(Pt 23):4170-81. doi: 10.1242/jcs.068387. Epub 2010 Nov 9. J Cell Sci. 2010. PMID: 21062895 Free PMC article.
-
New Aspects of the Contribution of ER to SOCE Regulation: TRPC Proteins as a Link Between Plasma Membrane Ion Transport and Intracellular Ca2+ Stores.Adv Exp Med Biol. 2017;993:239-255. doi: 10.1007/978-3-319-57732-6_13. Adv Exp Med Biol. 2017. PMID: 28900918 Review.
-
Protein-protein interaction and functionTRPC channels.Pflugers Arch. 2005 Oct;451(1):116-24. doi: 10.1007/s00424-005-1442-2. Epub 2005 Jul 26. Pflugers Arch. 2005. PMID: 16044307 Review.
Cited by
-
Live-cell imaging of ER-PM contact architecture by a novel TIRFM approach reveals extension of junctions in response to store-operated Ca2+-entry.Sci Rep. 2016 Oct 19;6:35656. doi: 10.1038/srep35656. Sci Rep. 2016. PMID: 27759093 Free PMC article.
-
Differential gene expression from genome-wide microarray analyses distinguishes Lohmann Selected Leghorn and Lohmann Brown layers.PLoS One. 2012;7(10):e46787. doi: 10.1371/journal.pone.0046787. Epub 2012 Oct 8. PLoS One. 2012. PMID: 23056453 Free PMC article.
-
Ca2+ store depletion causes STIM1 to accumulate in ER regions closely associated with the plasma membrane.J Cell Biol. 2006 Sep 11;174(6):803-13. doi: 10.1083/jcb.200604014. J Cell Biol. 2006. PMID: 16966422 Free PMC article.
-
The Role of Lipids in CRAC Channel Function.Biomolecules. 2022 Feb 23;12(3):352. doi: 10.3390/biom12030352. Biomolecules. 2022. PMID: 35327543 Free PMC article. Review.
-
C-terminal domain of chromogranin B regulates intracellular calcium signaling.J Biol Chem. 2011 Dec 30;286(52):44888-96. doi: 10.1074/jbc.M111.251330. Epub 2011 Oct 20. J Biol Chem. 2011. PMID: 22016391 Free PMC article.
References
-
- Barrero, M.J., M. Montero, and J. Alvarez. 1997. Dynamics of [Ca2+] in the endoplasmic reticulum and cytoplasm of intact HeLa cells. A comparative study. J. Biol. Chem. 272:27694–27699. - PubMed
-
- Bastianutto, C., E. Clementi, F. Codazzi, P. Podini, F. De Giorgi, R. Rizzuto, J. Meldolesi, and T. Pozzan. 1995. Overexpression of calreticulin increases the Ca2+ capacity of rapidly exchanging Ca2+ stores and reveals aspects of their lumenal microenvironment and function. J. Cell Biol. 130:847–855. - PMC - PubMed
-
- Berridge, M.J., P. Lipp, and M.D. Bootman. 2000. The versatility and universality of calcium signaling. Nat. Rev. Mol. Cell Biol. 1:11–21. - PubMed
-
- Bootman, M.D., T.J. Collins, L. MacKenzie, H.L. Roderick, M.J. Berridge, and C.M. Peppiat. 2002. 2-aminoethoxydiphenyl borate (2-APB) is a reliable blocker of store-operated Ca2+ entry but an inconsistent inhibitor of InP3-induced Ca2+ release. FASEB J. 16:1145–1150. - PubMed
-
- Boulay, G., D.M. Brown, N. Qin, M. Jiang, A. Dietrich, M.X. Zhu, Z. Chen, M. Birnbaumer, K. Mikoshiba, and L. Birnbaumer. 1999. Modulation of calcium entry by polypeptides of the IP3R that bind TRP: evidence for roles of TRP and IP3R in store depletion-activated Ca2+ entry. Proc. Natl. Acad. Sci. USA. 96:14955–14960. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
