

Gene targeting reveals a widespread role for the high-mobility-group transcription factor Sox11 in tissue remodeling
- PMID: 15254231
- PMCID: PMC444853
- DOI: 10.1128/MCB.24.15.6635-6644.2004
Gene targeting reveals a widespread role for the high-mobility-group transcription factor Sox11 in tissue remodeling
Abstract
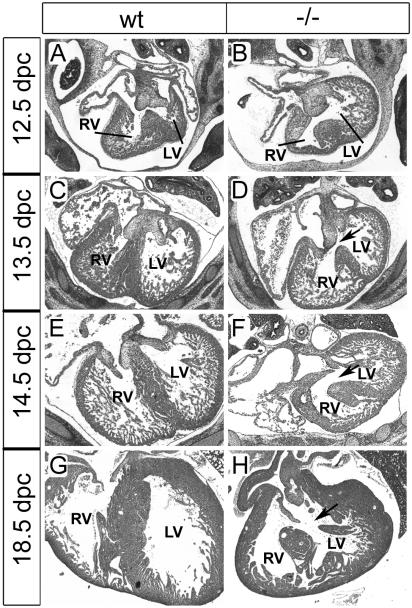
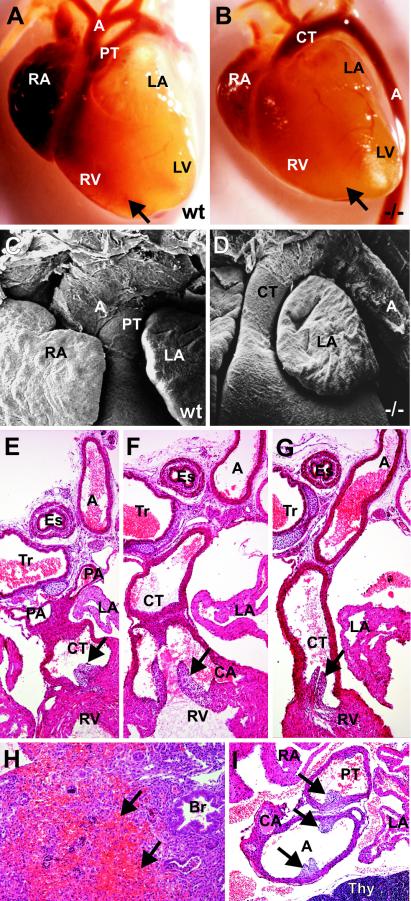
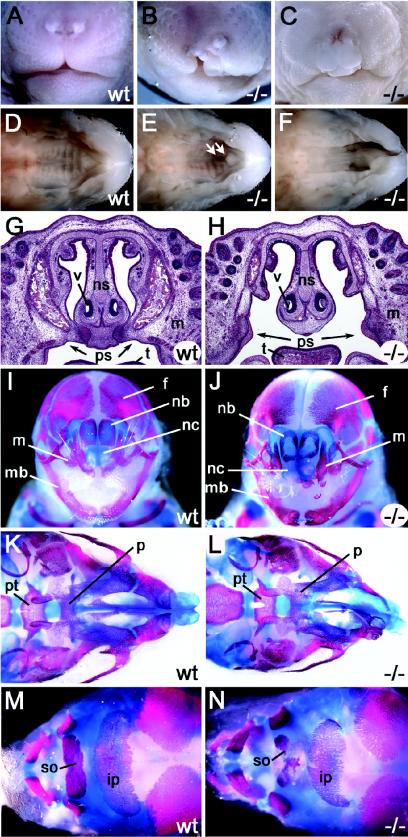
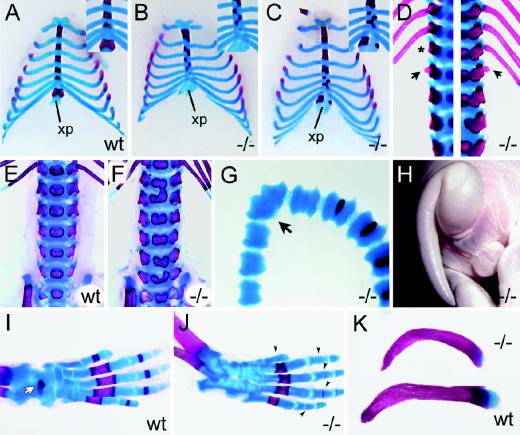
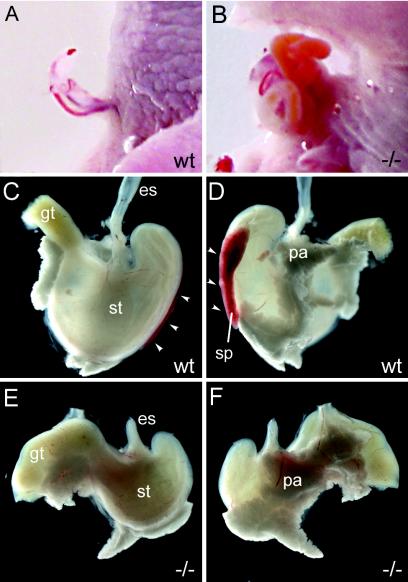
The high-mobility-group domain-containing transcription factor Sox11 is expressed transiently during embryonic development in many tissues that undergo inductive remodeling. Here we have analyzed the function of Sox11 by gene deletion in the mouse. Sox11-deficient mice died at birth from congenital cyanosis, likely resulting from heart defects. These included ventricular septation defects and outflow tract malformations that ranged from arterial common trunk to a condition known as double outlet right ventricle. Many other organs that normally express Sox11 also exhibited severe developmental defects. We observed various craniofacial and skeletal malformations, asplenia, and hypoplasia of the lung, stomach, and pancreas. Eyelids and the abdominal wall did not close properly in some Sox11-deficient mice. This phenotype suggests a prime function for Sox11 in tissue remodeling and identifies SOX11 as a potentially mutated gene in corresponding human malformation syndromes.
Figures
Similar articles
-
Sox12 deletion in the mouse reveals nonreciprocal redundancy with the related Sox4 and Sox11 transcription factors.Mol Cell Biol. 2008 Aug;28(15):4675-87. doi: 10.1128/MCB.00338-08. Epub 2008 May 27. Mol Cell Biol. 2008. PMID: 18505825 Free PMC article.
-
Foxp1 regulates cardiac outflow tract, endocardial cushion morphogenesis and myocyte proliferation and maturation.Development. 2004 Sep;131(18):4477-87. doi: 10.1242/dev.01287. Development. 2004. PMID: 15342473
-
Expression of Sox transcription factors in the developing mouse pancreas.Dev Dyn. 2003 Jul;227(3):402-8. doi: 10.1002/dvdy.10311. Dev Dyn. 2003. PMID: 12815626
-
Expression of the Sox11 gene in mouse embryos suggests roles in neuronal maturation and epithelio-mesenchymal induction.Dev Dyn. 1997 Oct;210(2):79-86. doi: 10.1002/(SICI)1097-0177(199710)210:2<79::AID-AJA1>3.0.CO;2-6. Dev Dyn. 1997. PMID: 9337129
-
Role of the vascular endothelial growth factor isoforms in retinal angiogenesis and DiGeorge syndrome.Verh K Acad Geneeskd Belg. 2005;67(4):229-76. Verh K Acad Geneeskd Belg. 2005. PMID: 16334858 Review.
Cited by
-
Transcription factors SOX4 and SOX11 function redundantly to regulate the development of mouse retinal ganglion cells.J Biol Chem. 2013 Jun 21;288(25):18429-38. doi: 10.1074/jbc.M113.478503. Epub 2013 May 6. J Biol Chem. 2013. PMID: 23649630 Free PMC article.
-
The role of tumor suppressor gene SOX11 in prostate cancer.Tumour Biol. 2015 Aug;36(8):6133-8. doi: 10.1007/s13277-015-3296-3. Epub 2015 Mar 14. Tumour Biol. 2015. PMID: 25773392
-
Aberrant SOX11 promoter methylation is associated with poor prognosis in gastric cancer.Cell Oncol (Dordr). 2015 Jun;38(3):183-94. doi: 10.1007/s13402-015-0219-7. Epub 2015 Mar 24. Cell Oncol (Dordr). 2015. PMID: 25801783
-
Clinicopathological features and prognostic value of SOX11 in childhood acute lymphoblastic leukemia.Sci Rep. 2020 Feb 6;10(1):2043. doi: 10.1038/s41598-020-58970-z. Sci Rep. 2020. PMID: 32029838 Free PMC article.
-
Integrative microRNA and proteomic approaches identify novel osteoarthritis genes and their collaborative metabolic and inflammatory networks.PLoS One. 2008;3(11):e3740. doi: 10.1371/journal.pone.0003740. Epub 2008 Nov 17. PLoS One. 2008. PMID: 19011694 Free PMC article.
References
-
- Abu-Issa, R., G. Smyth, I. Smoak, K.-I. Yamamura, and E. N. Meyera. 2002. Fgf8 is required for pharyngeal arch and cardiovascular development in the mouse. Development 129:4613-4625. - PubMed
-
- Bernstein, E. 2004. The cardiovascular system, p. 413-437. In R. E. Behrman, R. M. Kliegman, and A. M. Arvin (ed.), Nelson textbook of pediatrics, 17th ed. Saunders, Philadelphia, Pa.
-
- Bowles, J., G. Schepers, and P. Koopman. 2000. Phylogeny of the SOX family of developmental transcription factors based on sequence and structural indicators. Dev. Biol. 227:239-255. - PubMed
-
- Brewer, S., W. Feng, J. Huang, S. Sullivan, and T. Williams. 2004. Wnt1-Cre-mediated deletion of AP-2alpha causes multiple neural crest-related defects. Dev. Biol. 267:132-152. - PubMed
-
- Brewer, S., X. Jiang, S. Donaldson, T. Williams, and H. M. Sucov. 2002. Requirement for AP-2α in cardiac outflow tract morphogenesis. Mech. Dev. 110:139-149. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases