

Functional analysis of the Streptococcus gordonii DL1 sialic acid-binding adhesin and its essential role in bacterial binding to platelets
- PMID: 15213130
- PMCID: PMC427394
- DOI: 10.1128/IAI.72.7.3876-3882.2004
Functional analysis of the Streptococcus gordonii DL1 sialic acid-binding adhesin and its essential role in bacterial binding to platelets
Abstract
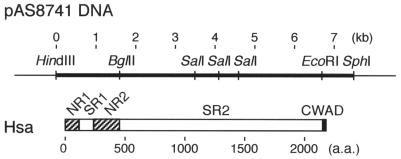
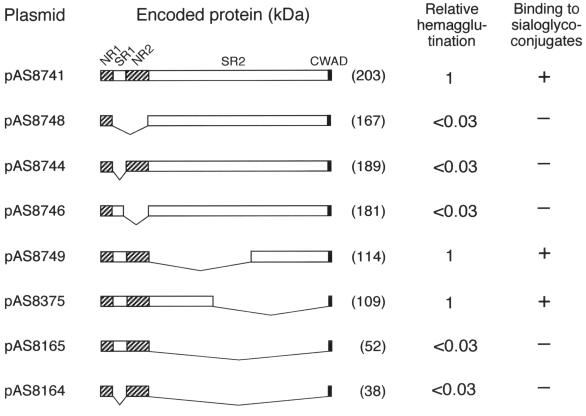
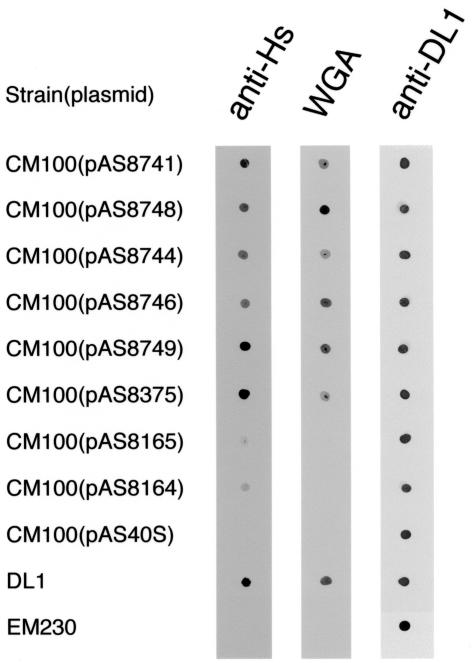
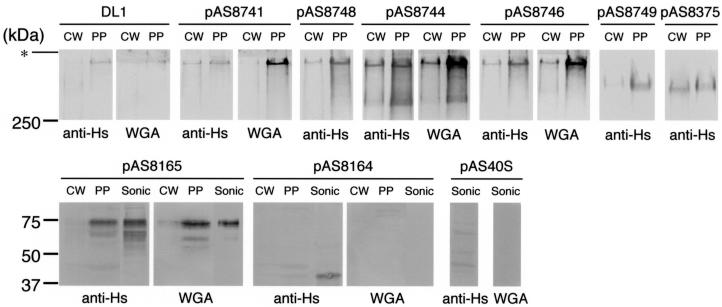
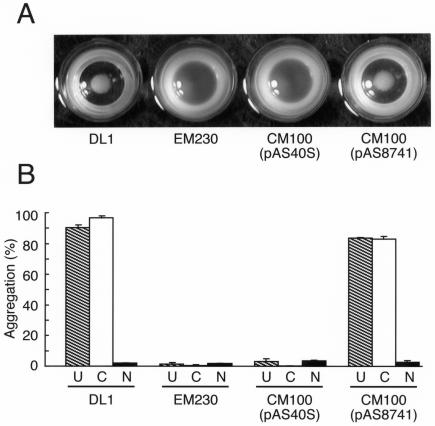
Bacterial recognition of host sialic acid-containing receptors plays an important role in microbial colonization of the human oral cavity. The sialic acid-binding adhesin of Streptococcus gordonii DL1 was previously associated with the hsa gene encoding a 203-kDa protein. The predicted protein sequence consists of an N-terminal nonrepetitive region (NR1), including a signal sequence, a relatively short serine-rich region (SR1), a second nonrepetitive region (NR2), a long serine-rich region (SR2) containing 113 dodecapeptide repeats, and a C-terminal cell wall anchoring domain. In the present study, the contributions of SR1, NR2, and SR2 to Hsa-mediated adhesion were assessed by genetic complementation. Adhesion of an hsa chromosomal deletion mutant to sialic acid-containing receptors was restored by plasmids containing hsa constructs encoding Hsa that lacked either the N- or C-terminal portion of SR2. In contrast, hsa constructs that lacked the coding sequences for SR1, NR2, or the entire SR2 region failed to restore adhesion. Surface expression of recombinant Hsa was not affected by removal of SR1, NR2, or a portion of SR2 but was greatly reduced by complete removal of SR2. Wheat germ agglutinin, a probe for Hsa-specific glycosylation, reacted with recombinant Hsa lacking SR1, NR2, or SR2 but not with recombinant Hsa lacking both SR1 and SR2. Significantly, the aggregation of human platelets by S. gordonii DL1, an interaction implicated in the pathogenesis of infective endocarditis, required the expression of hsa. Moreover, neuraminidase treatment of the platelets eliminated this interaction, further supporting the hypothesis that Hsa plays an essential role in the bacterium-platelet interaction.
Figures
Similar articles
-
Hsa, an adhesin of Streptococcus gordonii DL1, binds to alpha2-3-linked sialic acid on glycophorin A of the erythrocyte membrane.Microbiol Immunol. 2008 Feb;52(2):69-77. doi: 10.1111/j.1348-0421.2008.00015.x. Microbiol Immunol. 2008. PMID: 18380804
-
Identification and characterization of hsa, the gene encoding the sialic acid-binding adhesin of Streptococcus gordonii DL1.Infect Immun. 2002 Mar;70(3):1209-18. doi: 10.1128/IAI.70.3.1209-1218.2002. Infect Immun. 2002. PMID: 11854202 Free PMC article.
-
Two Arginine Residues of Streptococcus gordonii Sialic Acid-Binding Adhesin Hsa Are Essential for Interaction to Host Cell Receptors.PLoS One. 2016 Apr 21;11(4):e0154098. doi: 10.1371/journal.pone.0154098. eCollection 2016. PLoS One. 2016. PMID: 27101147 Free PMC article.
-
[Adhesins of oral streptococci].Nihon Saikingaku Zasshi. 2013;68(2):283-93. doi: 10.3412/jsb.68.283. Nihon Saikingaku Zasshi. 2013. PMID: 23727707 Review. Japanese.
-
A role for glycosylated serine-rich repeat proteins in gram-positive bacterial pathogenesis.Mol Oral Microbiol. 2012 Aug;27(4):257-69. doi: 10.1111/j.2041-1014.2012.00653.x. Epub 2012 Jun 11. Mol Oral Microbiol. 2012. PMID: 22759311 Free PMC article. Review.
Cited by
-
Serine-rich repeat proteins from gut microbes.Gut Microbes. 2020;11(1):102-117. doi: 10.1080/19490976.2019.1602428. Epub 2019 Apr 29. Gut Microbes. 2020. PMID: 31035824 Free PMC article. Review.
-
Characterization of exosome-like vesicles released from human tracheobronchial ciliated epithelium: a possible role in innate defense.FASEB J. 2009 Jun;23(6):1858-68. doi: 10.1096/fj.08-119131. Epub 2009 Feb 3. FASEB J. 2009. PMID: 19190083 Free PMC article.
-
Multiple adhesin proteins on the cell surface of Streptococcus gordonii are involved in adhesion to human fibronectin.Microbiology (Reading). 2009 Nov;155(Pt 11):3572-3580. doi: 10.1099/mic.0.032078-0. Epub 2009 Aug 6. Microbiology (Reading). 2009. PMID: 19661180 Free PMC article.
-
Inactivation of Streptococcus gordonii SspAB alters expression of multiple adhesin genes.Infect Immun. 2005 Jun;73(6):3351-7. doi: 10.1128/IAI.73.6.3351-3357.2005. Infect Immun. 2005. PMID: 15908361 Free PMC article.
-
Alteration of the neuronal and glial cell profiles in Neu1-deficient zebrafish.Glycoconj J. 2022 Aug;39(4):499-512. doi: 10.1007/s10719-022-10074-8. Epub 2022 Jul 25. Glycoconj J. 2022. PMID: 35877057
References
-
- Bensing, B. A., and P. M. Sullam. 2002. An accessory sec locus of Streptococcus gordonii is required for export of the surface protein GspB and for normal levels of binding to human platelets. Mol. Microbiol. 44:1081-1094. - PubMed
-
- Demuth, D. R., Y. Duan, W. Brooks, A. R. Holmes, R. McNab, and H. F. Jenkinson. 1996. Tandem genes encode cell-surface polypeptides SspA and SspB which mediate adhesion of the oral bacterium Streptococcus gordonii to human and bacterial receptors. Mol. Microbiol. 20:403-413. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
