

Promoter unwinding and promoter clearance by RNA polymerase: detection by single-molecule DNA nanomanipulation
- PMID: 15037753
- PMCID: PMC387324
- DOI: 10.1073/pnas.0307241101
Promoter unwinding and promoter clearance by RNA polymerase: detection by single-molecule DNA nanomanipulation
Abstract
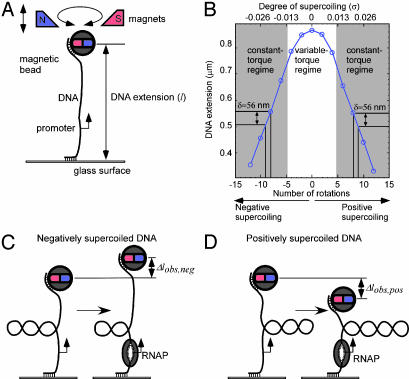
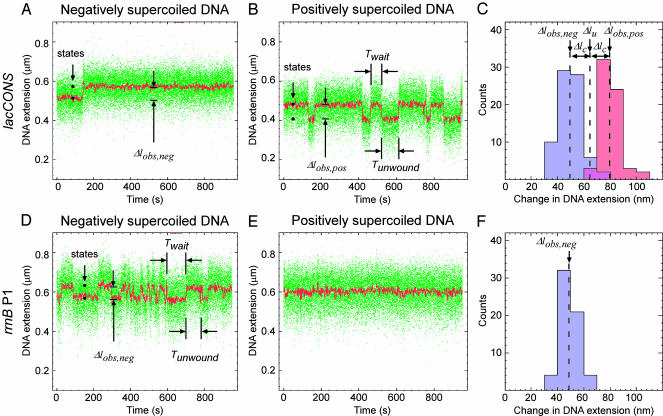
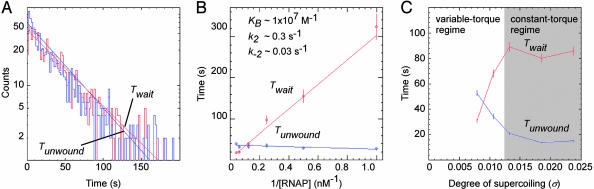
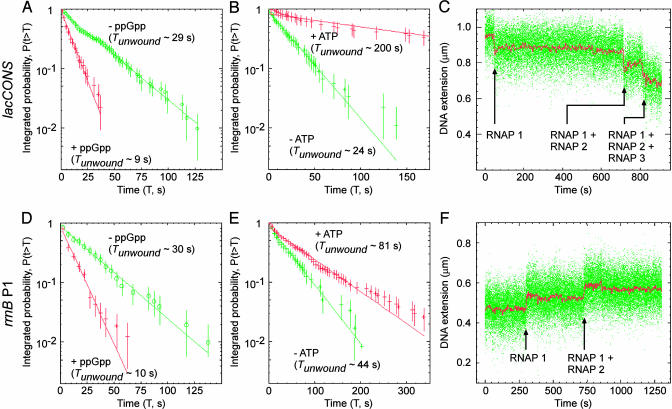
By monitoring the end-to-end extension of a mechanically stretched, supercoiled, single DNA molecule, we have been able directly to observe the change in extension associated with unwinding of approximately one turn of promoter DNA by RNA polymerase (RNAP). By performing parallel experiments with negatively and positively supercoiled DNA, we have been able to deconvolute the change in extension caused by RNAP-dependent DNA unwinding (with approximately 1-bp resolution) and the change in extension caused by RNAP-dependent DNA compaction (with approximately 5-nm resolution). We have used this approach to quantify the extent of unwinding and compaction, the kinetics of unwinding and compaction, and effects of supercoiling, sequence, ppGpp, and nucleotides. We also have used this approach to detect promoter clearance and promoter recycling by successive RNAP molecules. We find that the rate of formation and the stability of the unwound complex depend profoundly on supercoiling and that supercoiling exerts its effects mechanically (through torque), and not structurally (through the number and position of supercoils). The approach should permit analysis of other nucleic-acid-processing factors that cause changes in DNA twist and/or DNA compaction.
Figures
Similar articles
-
Single-molecule visualization of twin-supercoiled domains generated during transcription.Nucleic Acids Res. 2024 Feb 28;52(4):1677-1687. doi: 10.1093/nar/gkad1181. Nucleic Acids Res. 2024. PMID: 38084930 Free PMC article.
-
Single-molecule DNA nanomanipulation: detection of promoter-unwinding events by RNA polymerase.Methods Enzymol. 2003;370:577-98. doi: 10.1016/S0076-6879(03)70049-4. Methods Enzymol. 2003. PMID: 14712677 No abstract available.
-
Transcription initiation at a consensus bacterial promoter proceeds via a 'bind-unwind-load-and-lock' mechanism.Elife. 2021 Oct 11;10:e70090. doi: 10.7554/eLife.70090. Elife. 2021. PMID: 34633286 Free PMC article.
-
The Role of Supercoiling in the Motor Activity of RNA Polymerases.Methods Mol Biol. 2018;1805:215-232. doi: 10.1007/978-1-4939-8556-2_11. Methods Mol Biol. 2018. PMID: 29971720 Review.
-
Single-molecule analysis of RNA polymerase transcription.Annu Rev Biophys Biomol Struct. 2006;35:343-60. doi: 10.1146/annurev.biophys.35.010406.150153. Annu Rev Biophys Biomol Struct. 2006. PMID: 16689640 Review.
Cited by
-
Topoisomerase IIα represses transcription by enforcing promoter-proximal pausing.Cell Rep. 2021 Apr 13;35(2):108977. doi: 10.1016/j.celrep.2021.108977. Cell Rep. 2021. PMID: 33852840 Free PMC article.
-
Transcription-dependent dynamic supercoiling is a short-range genomic force.Nat Struct Mol Biol. 2013 Mar;20(3):396-403. doi: 10.1038/nsmb.2517. Epub 2013 Feb 17. Nat Struct Mol Biol. 2013. PMID: 23416947 Free PMC article.
-
Interaction of Escherichia coli RNA polymerase σ70 subunit with promoter elements in the context of free σ70, RNA polymerase holoenzyme, and the β'-σ70 complex.J Biol Chem. 2011 Jan 7;286(1):270-9. doi: 10.1074/jbc.M110.174102. Epub 2010 Oct 15. J Biol Chem. 2011. PMID: 20952386 Free PMC article.
-
Single-molecule studies of RNA polymerase: motoring along.Annu Rev Biochem. 2008;77:149-76. doi: 10.1146/annurev.biochem.77.073106.100741. Annu Rev Biochem. 2008. PMID: 18410247 Free PMC article. Review.
-
Transcriptional regulation in Saccharomyces cerevisiae: transcription factor regulation and function, mechanisms of initiation, and roles of activators and coactivators.Genetics. 2011 Nov;189(3):705-36. doi: 10.1534/genetics.111.127019. Genetics. 2011. PMID: 22084422 Free PMC article. Review.
References
-
- Record, M. T. J., Reznikoff, W., Craig, M., McQuade, K. & Schlax, P. (1996) in Escherichia coli and Salmonella, ed. Neidhart, F. C. (Am. Soc. Microbiol. Press, Washington, DC), Vol. 1, pp. 792–820.
-
- Strick, T., Allemand, J., Bensimon, D., Bensimon, A. & Croquette, V. (1996) Science 271, 1835–1837. - PubMed
-
- Revyakin, A., Allemand, J.-F., Croquette, V., Ebright, R. & Strick, T. (2003) Methods Enzymol. 370, 577–598. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
