

The SNARE Ykt6 mediates protein palmitoylation during an early stage of homotypic vacuole fusion
- PMID: 14685280
- PMCID: PMC1271655
- DOI: 10.1038/sj.emboj.7600015
The SNARE Ykt6 mediates protein palmitoylation during an early stage of homotypic vacuole fusion
Abstract
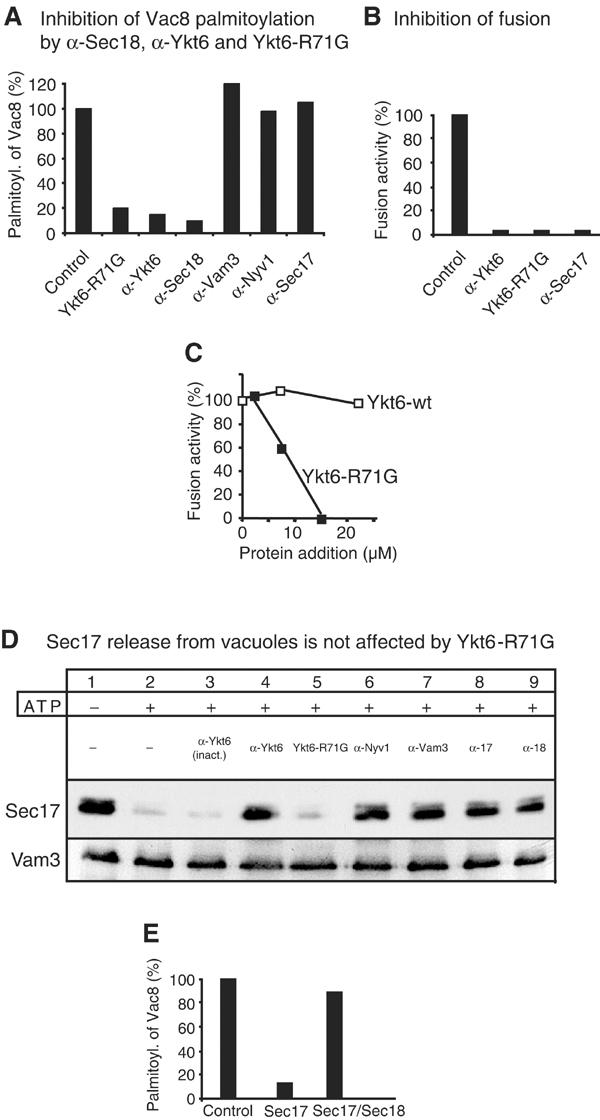
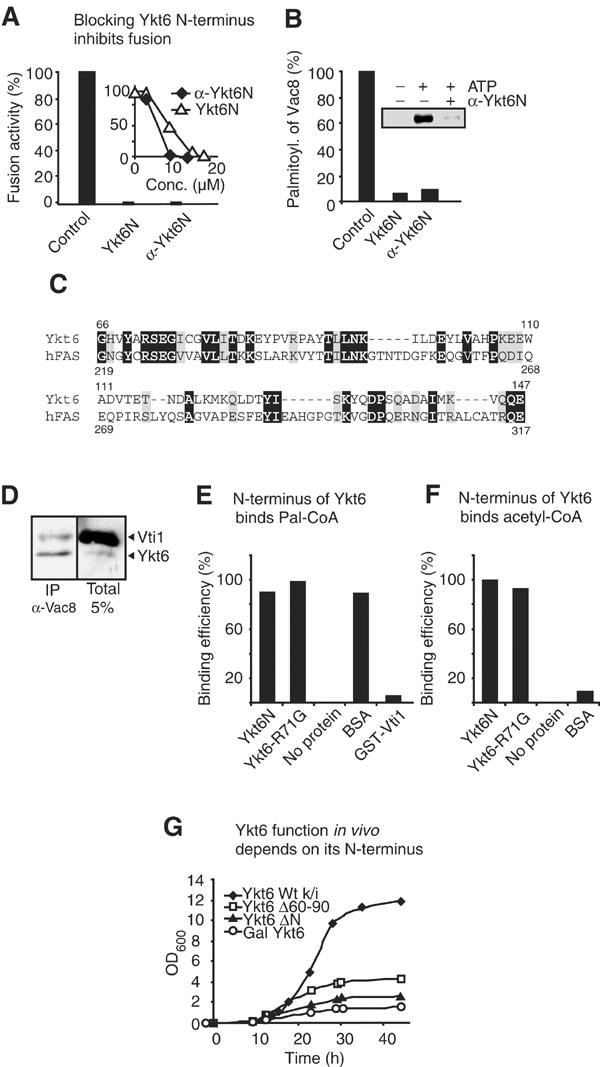
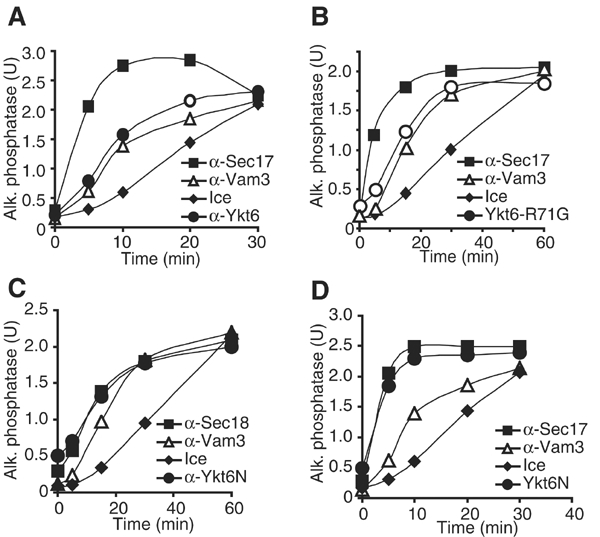
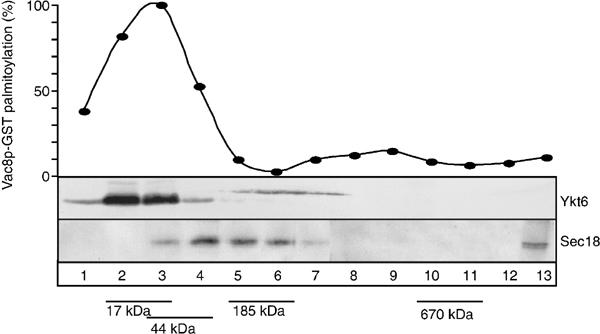
The NSF homolog Sec18 initiates fusion of yeast vacuoles by disassembling cis-SNARE complexes during priming. Sec18 is also required for palmitoylation of the fusion factor Vac8, although the acylation machinery has not been identified. Here we show that the SNARE Ykt6 mediates Vac8 palmitoylation and acts during a novel subreaction of vacuole fusion. This subreaction is controlled by a Sec17-independent function of Sec18. Our data indicate that Ykt6 presents Pal-CoA via its N-terminal longin domain to Vac8, while transfer to Vac8's SH4 domain occurs spontaneously and not enzymatically. The conservation of Ykt6 and its localization to several organelles suggest that its acyltransferase activity may also be required in other intracellular fusion events.
Figures
Similar articles
-
ATP-independent control of Vac8 palmitoylation by a SNARE subcomplex on yeast vacuoles.J Biol Chem. 2005 Apr 15;280(15):15348-55. doi: 10.1074/jbc.M410582200. Epub 2005 Feb 8. J Biol Chem. 2005. PMID: 15701652
-
Vac8p release from the SNARE complex and its palmitoylation are coupled and essential for vacuole fusion.EMBO J. 2001 Jun 15;20(12):3145-55. doi: 10.1093/emboj/20.12.3145. EMBO J. 2001. PMID: 11406591 Free PMC article.
-
The SNARE Ykt6 is released from yeast vacuoles during an early stage of fusion.EMBO Rep. 2005 Mar;6(3):245-50. doi: 10.1038/sj.embor.7400350. EMBO Rep. 2005. PMID: 15723044 Free PMC article.
-
Probing protein palmitoylation at the yeast vacuole.Methods. 2006 Oct;40(2):171-6. doi: 10.1016/j.ymeth.2006.06.020. Methods. 2006. PMID: 17012029 Review.
-
Structure and function of longin SNAREs.J Cell Sci. 2015 Dec 1;128(23):4263-72. doi: 10.1242/jcs.178574. Epub 2015 Nov 13. J Cell Sci. 2015. PMID: 26567219 Review.
Cited by
-
Localization and activity of the SNARE Ykt6 determined by its regulatory domain and palmitoylation.Proc Natl Acad Sci U S A. 2004 Apr 6;101(14):4815-20. doi: 10.1073/pnas.0401183101. Epub 2004 Mar 24. Proc Natl Acad Sci U S A. 2004. PMID: 15044687 Free PMC article.
-
LncRNA PVT1 promotes exosome secretion through YKT6, RAB7, and VAMP3 in pancreatic cancer.Aging (Albany NY). 2020 Jun 4;12(11):10427-10440. doi: 10.18632/aging.103268. Epub 2020 Jun 4. Aging (Albany NY). 2020. PMID: 32499447 Free PMC article.
-
The multi-functional SNARE protein Ykt6 in autophagosomal fusion processes.Cell Cycle. 2019 Mar-Apr;18(6-7):639-651. doi: 10.1080/15384101.2019.1580488. Epub 2019 Mar 17. Cell Cycle. 2019. PMID: 30836834 Free PMC article. Review.
-
Non-canonical role of the SNARE protein Ykt6 in autophagosome-lysosome fusion.PLoS Genet. 2018 Apr 25;14(4):e1007359. doi: 10.1371/journal.pgen.1007359. eCollection 2018 Apr. PLoS Genet. 2018. PMID: 29694367 Free PMC article.
-
Global analysis of protein palmitoylation in yeast.Cell. 2006 Jun 2;125(5):1003-13. doi: 10.1016/j.cell.2006.03.042. Cell. 2006. PMID: 16751107 Free PMC article.
References
-
- Antonin W, Fasshauer D, Becker S, Jahn R, Schneider TR (2002) Crystal structure of the endosomal SNARE complex reveals common structural principles of all SNAREs. Nat Struct Biol 9: 107–111 - PubMed
-
- Bizzozero OA, Bixler HA, Pastuszyn A (2001) Structural determinants influencing the reaction of cysteine-containing peptides with palmitoyl-coenzyme A and other thioesters. Biochem Biophys Acta 1545: 278–288 - PubMed
-
- Catchpoole DR, Hong W (1999) Characterization of the sequence and expression of a Ykt6p prenylated SNARE from rat. DNA Cell Biol 18: 141–145 - PubMed
-
- Chen YA, Scheller RH (2001) SNARE-mediated membrane fusion. Nat Rev Mol Cell Biol 2: 98–106 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
