

The membrane-proximal region of vesicular stomatitis virus glycoprotein G ectodomain is critical for fusion and virus infectivity
- PMID: 14610202
- PMCID: PMC262588
- DOI: 10.1128/jvi.77.23.12807-12818.2003
The membrane-proximal region of vesicular stomatitis virus glycoprotein G ectodomain is critical for fusion and virus infectivity
Abstract
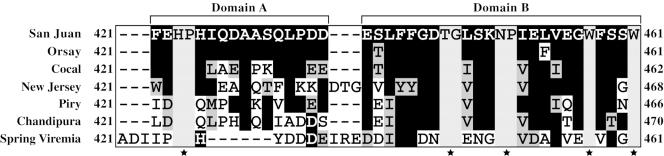
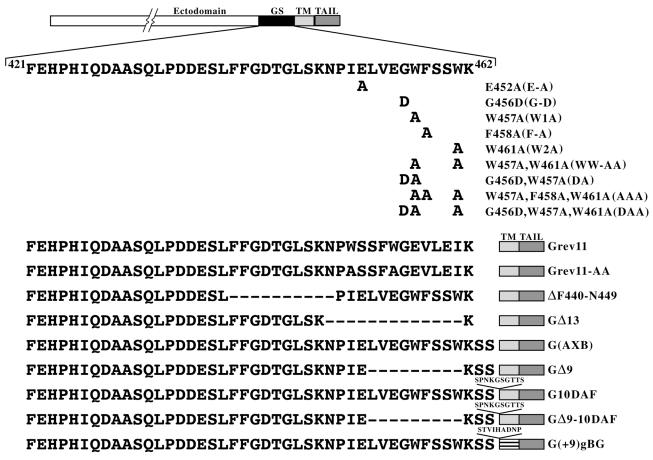
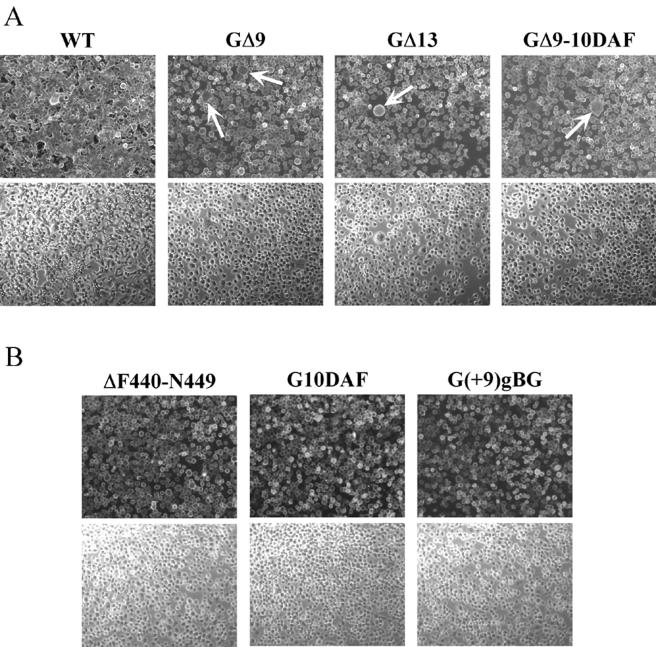
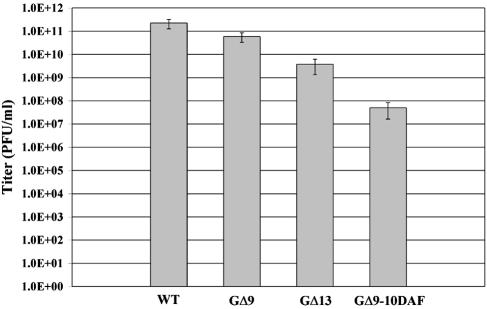
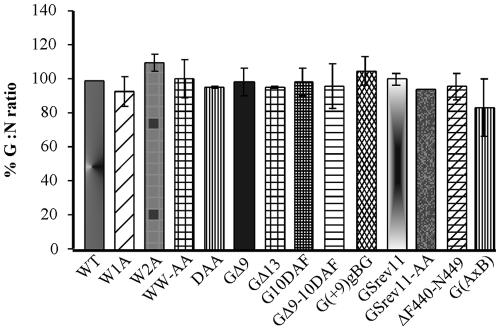
The glycoprotein (G) of vesicular stomatitis virus (VSV) is responsible for binding of virus to cells and for mediating virus entry following endocytosis by inducing fusion of the viral envelope with the endosomal membrane. The fusion peptide of G is internal (residues 116 to 137) and exhibits characteristics similar to those of other internal fusion peptides, but recent studies have implicated the region adjacent to the transmembrane domain as also being important for G-mediated membrane fusion. Sequence alignment of the membrane-proximal region of G from several different vesiculoviruses revealed that this domain is highly conserved, suggesting that it is important for G function. Mutational analysis was used to show that this region is not essential for G protein oligomerization, transport to the cell surface, or incorporation into virus particles but that it is essential for acid-induced membrane fusion activity and for virus infectivity. Deletion of the 13 membrane-proximal amino acids (N449 to W461) dramatically reduced cell-cell fusion activity and reduced virus infectivity approximately 100-fold, but mutation of conserved aromatic residues (W457, F458, and W461) either singly or together had only modest effects on cell-cell fusion activity; recombinant virus encoding these mutants replicated as efficiently as wild-type (WT) VSV. Insertion of heterologous sequences in the juxtamembrane region completely abolished membrane fusion activity and virus infectivity, as did deletion of residues F440 to N449. The insertion mutants showed some changes in pH-dependent conformational changes and in virus binding, which could partially explain the defects in membrane fusion activity, but all the other mutants were similar to WT G with respect to conformational changes and virus binding. These data support the hypothesis that the membrane-proximal domain contributes to G-mediated membrane fusion activity, yet the conserved aromatic residues are not essential for membrane fusion or virus infectivity.
Figures
Similar articles
-
The membrane-proximal domain of vesicular stomatitis virus G protein functions as a membrane fusion potentiator and can induce hemifusion.J Virol. 2002 Dec;76(23):12300-11. doi: 10.1128/jvi.76.23.12300-12311.2002. J Virol. 2002. PMID: 12414970 Free PMC article.
-
Vesicular stomatitis virus glycoprotein mutations that affect membrane fusion activity and abolish virus infectivity.J Virol. 1995 Mar;69(3):1435-43. doi: 10.1128/JVI.69.3.1435-1443.1995. J Virol. 1995. PMID: 7853475 Free PMC article.
-
Attenuation of recombinant vesicular stomatitis viruses encoding mutant glycoproteins demonstrate a critical role for maintaining a high pH threshold for membrane fusion in viral fitness.Virology. 1998 Jan 20;240(2):349-58. doi: 10.1006/viro.1997.8921. Virology. 1998. PMID: 9454708
-
Structures of vesicular stomatitis virus glycoprotein: membrane fusion revisited.Cell Mol Life Sci. 2008 Jun;65(11):1716-28. doi: 10.1007/s00018-008-7534-3. Cell Mol Life Sci. 2008. PMID: 18345480 Free PMC article. Review.
-
Viral inactivation based on inhibition of membrane fusion: understanding the role of histidine protonation to develop new viral vaccines.Protein Pept Lett. 2009;16(7):779-85. doi: 10.2174/092986609788681823. Protein Pept Lett. 2009. PMID: 19601907 Review.
Cited by
-
Hydrophobic residues that form putative fusion loops of Epstein-Barr virus glycoprotein B are critical for fusion activity.J Virol. 2007 Sep;81(17):9596-600. doi: 10.1128/JVI.00758-07. Epub 2007 Jun 6. J Virol. 2007. PMID: 17553877 Free PMC article.
-
The structure of herpesvirus fusion glycoprotein B-bilayer complex reveals the protein-membrane and lateral protein-protein interaction.Structure. 2013 Aug 6;21(8):1396-405. doi: 10.1016/j.str.2013.05.018. Epub 2013 Jul 11. Structure. 2013. PMID: 23850455 Free PMC article.
-
Entry of rhabdoviruses into animal cells.Adv Exp Med Biol. 2013;790:167-77. doi: 10.1007/978-1-4614-7651-1_9. Adv Exp Med Biol. 2013. PMID: 23884591 Free PMC article. Review.
-
Replication-competent recombinant vesicular stomatitis virus encoding hepatitis C virus envelope proteins.J Virol. 2007 Aug;81(16):8601-12. doi: 10.1128/JVI.00608-07. Epub 2007 Jun 6. J Virol. 2007. PMID: 17553880 Free PMC article.
-
Neurovirulence properties of recombinant vesicular stomatitis virus vectors in non-human primates.Virology. 2007 Mar 30;360(1):36-49. doi: 10.1016/j.virol.2006.10.026. Epub 2006 Nov 13. Virology. 2007. PMID: 17098273 Free PMC article.
References
-
- Baker, K. A., R. E. Dutch, R. A. Lamb, and T. S. Jardetzky. 1999. Structural basis for paramyxovirus-mediated membrane fusion. Mol. Cell 3:309-319. - PubMed
-
- Bhella, R. S., S. T. Nichol, E. Wanas, and H. P. Ghosh. 1998. Structure, expression and phylogenetic analysis of the glycoprotein gene of Cocal virus. Virus Res. 54:197-205. - PubMed
-
- Bjorklund, H. V., K. H. Higman, and G. Kurath. 1996. The glycoprotein genes and gene junctions of the fish rhabdoviruses spring viremia of carp virus and hirame rhabdovirus: analysis of relationships with other rhabdoviruses. Virus Res. 42:65-80. - PubMed
-
- Brun, G., X. Bao, and L. Prevec. 1995. The relationship of Piry virus to other vesiculoviruses: a re-evaluation based on the glycoprotein gene sequence. Intervirology 38:274-282. - PubMed
-
- Bullough, P. A., F. M. Hughson, J. J. Skehel, and D. C. Wiley. 1994. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature 371:37-43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
