

Mitochondrial translocation contact sites: separation of dynamic and stabilizing elements in formation of a TOM-TIM-preprotein supercomplex
- PMID: 14532110
- PMCID: PMC213786
- DOI: 10.1093/emboj/cdg532
Mitochondrial translocation contact sites: separation of dynamic and stabilizing elements in formation of a TOM-TIM-preprotein supercomplex
Abstract
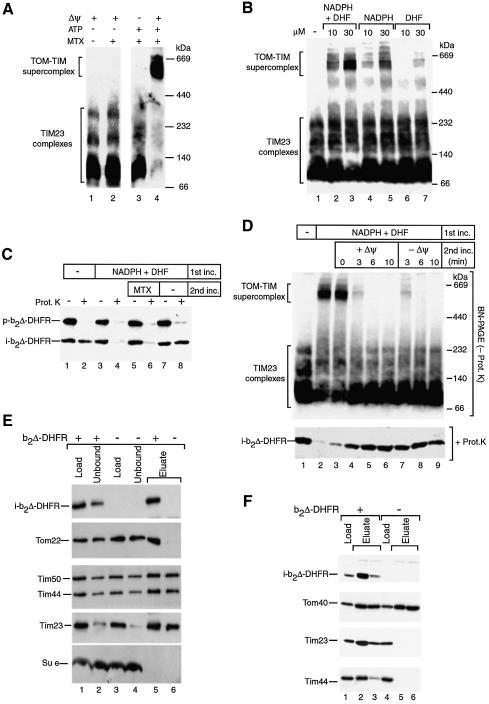
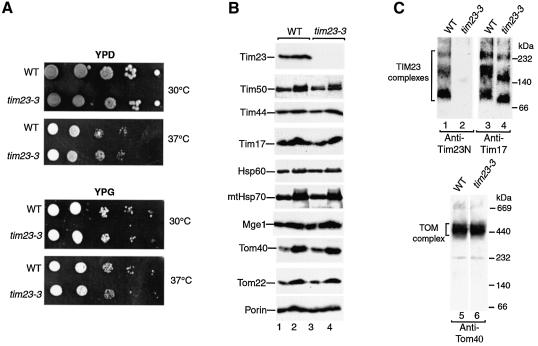
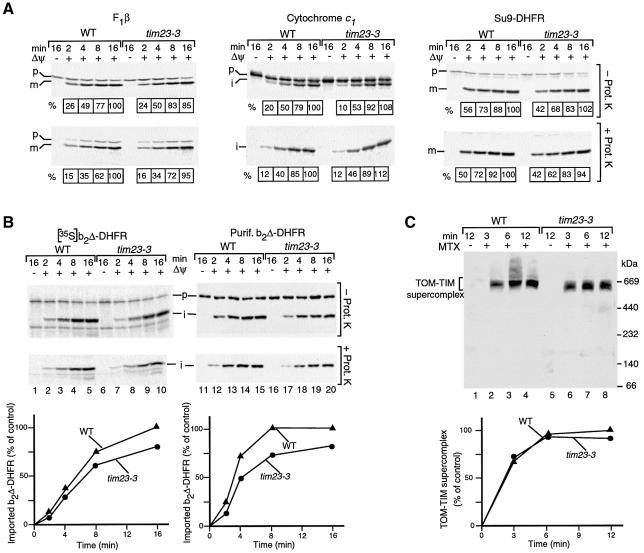
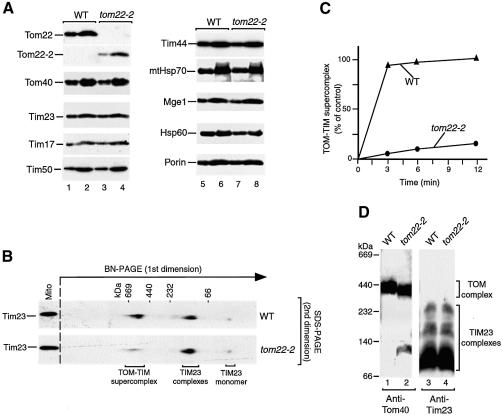
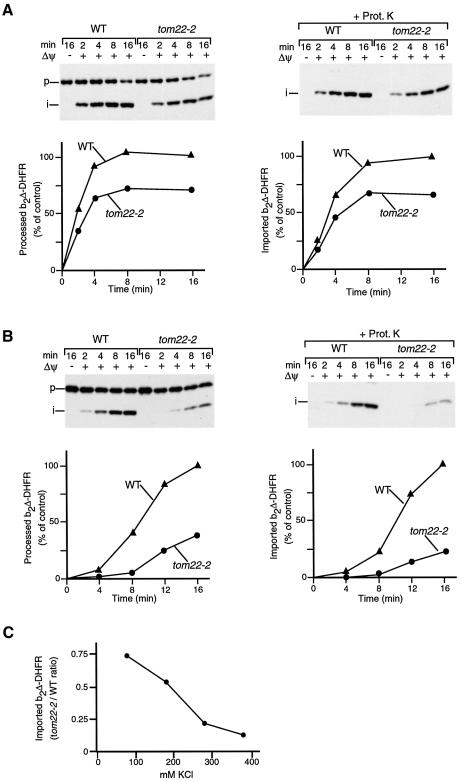
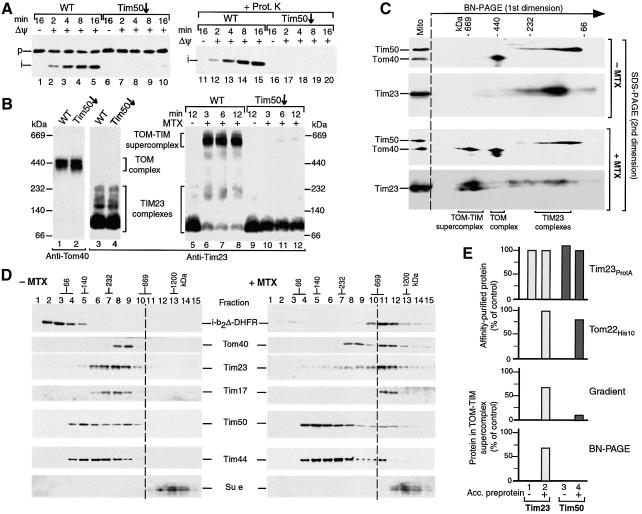
Preproteins with N-terminal presequences are imported into mitochondria at translocation contact sites that include the translocase of the outer membrane (TOM complex) and the presequence translocase of the inner membrane (TIM23 complex). Little is known about the functional cooperation of these translocases. We have characterized translocation contact sites by a productive TOM-TIM-preprotein supercomplex to address the role of three translocase subunits that expose domains to the intermembrane space (IMS). The IMS domain of the receptor Tom22 is required for stabilization of the translocation contact site supercomplex. Surprisingly, the N-terminal segment of the channel Tim23, which tethers the TIM23 complex to the outer membrane, is dispensable for both protein import and generation of the TOM-TIM supercomplex. Tim50, with its large IMS domain, is crucial for generation but not for stabilization of the supercomplex. Thus, Tim50 functions as a dynamic factor and the IMS domain of Tom22 represents a stabilizing element in formation of a productive translocation contact site supercomplex.
Figures
Similar articles
-
A presequence- and voltage-sensitive channel of the mitochondrial preprotein translocase formed by Tim23.Nat Struct Biol. 2001 Dec;8(12):1074-82. doi: 10.1038/nsb726. Nat Struct Biol. 2001. PMID: 11713477
-
Mitochondrial presequence translocase: switching between TOM tethering and motor recruitment involves Tim21 and Tim17.Cell. 2005 Mar 25;120(6):817-29. doi: 10.1016/j.cell.2005.01.011. Cell. 2005. PMID: 15797382
-
Tom22 is a multifunctional organizer of the mitochondrial preprotein translocase.Nature. 1999 Sep 30;401(6752):485-9. doi: 10.1038/46802. Nature. 1999. PMID: 10519552
-
Protein import into mitochondria.IUBMB Life. 2001 Sep-Nov;52(3-5):101-12. doi: 10.1080/15216540152845894. IUBMB Life. 2001. PMID: 11798021 Review.
-
Mitochondrial preprotein translocases as dynamic molecular machines.FEMS Yeast Res. 2006 Sep;6(6):849-61. doi: 10.1111/j.1567-1364.2006.00134.x. FEMS Yeast Res. 2006. PMID: 16911507 Review.
Cited by
-
Interplay between Mitochondrial Protein Import and Respiratory Complexes Assembly in Neuronal Health and Degeneration.Life (Basel). 2021 May 11;11(5):432. doi: 10.3390/life11050432. Life (Basel). 2021. PMID: 34064758 Free PMC article. Review.
-
Mechanisms of protein sorting in mitochondria.Cold Spring Harb Perspect Biol. 2012 Oct 1;4(10):a011320. doi: 10.1101/cshperspect.a011320. Cold Spring Harb Perspect Biol. 2012. PMID: 23028120 Free PMC article. Review.
-
ROMO1 is a constituent of the human presequence translocase required for YME1L protease import.J Cell Biol. 2019 Feb 4;218(2):598-614. doi: 10.1083/jcb.201806093. Epub 2018 Dec 31. J Cell Biol. 2019. PMID: 30598479 Free PMC article.
-
Mapping protein interactions in the active TOM-TIM23 supercomplex.Nat Commun. 2021 Sep 29;12(1):5715. doi: 10.1038/s41467-021-26016-1. Nat Commun. 2021. PMID: 34588454 Free PMC article.
-
A two-step mitochondrial import pathway couples the disulfide relay with matrix complex I biogenesis.J Cell Biol. 2023 Jul 3;222(7):e202210019. doi: 10.1083/jcb.202210019. Epub 2023 May 9. J Cell Biol. 2023. PMID: 37159021 Free PMC article.
References
-
- Abe Y., Shodai,T., Muto,T., Mihara,K., Torii,H., Nishikawa,S., Endo,T. and Kohda,D. (2000) Structural basis of presequence recognition by the mitochondrial protein import receptor Tom20. Cell, 100, 551–560. - PubMed
-
- Bauer M.F., Sirrenberg,C., Neupert,W. and Brunner,M. (1996) Role of Tim23 as voltage sensor and presequence receptor in protein import into mitochondria. Cell, 87, 33–41. - PubMed
-
- Bauer M.F., Hofmann,S., Neupert,W. and Brunner,M. (2000) Protein translocation into mitochondria: the role of TIM complexes. Trends Cell Biol., 10, 25–31. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
