

Investigation of structural and functional motifs within the vaccinia virus A14 phosphoprotein, an essential component of the virion membrane
- PMID: 12885904
- PMCID: PMC167248
- DOI: 10.1128/jvi.77.16.8857-8871.2003
Investigation of structural and functional motifs within the vaccinia virus A14 phosphoprotein, an essential component of the virion membrane
Abstract
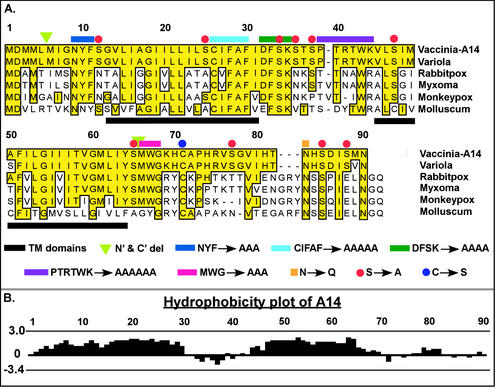
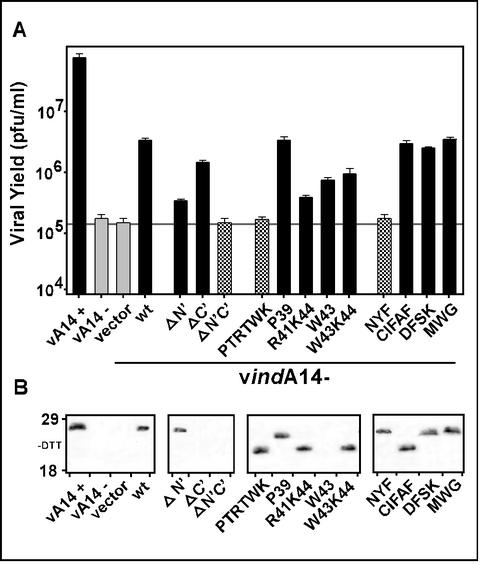
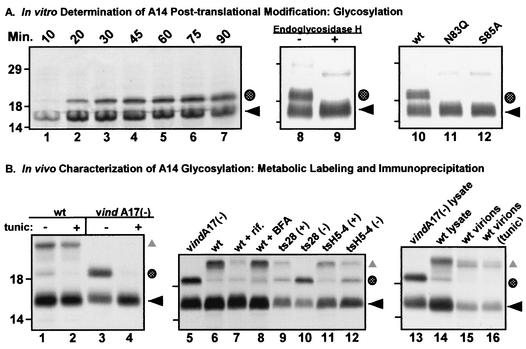
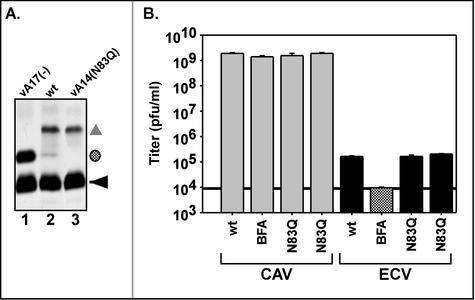
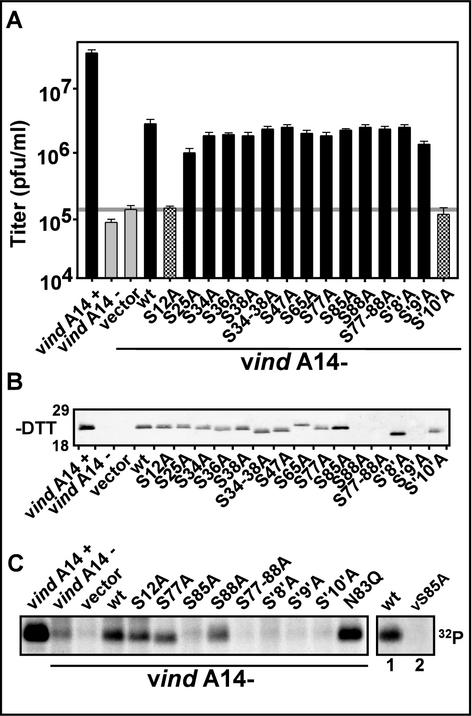
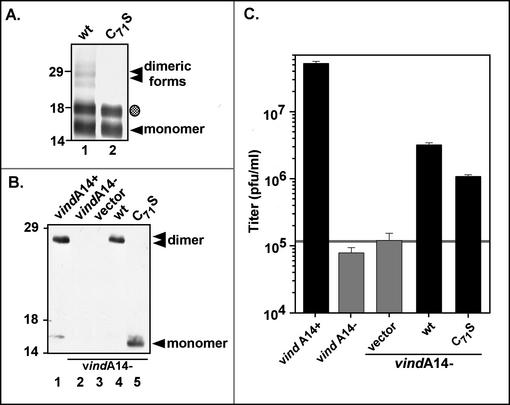
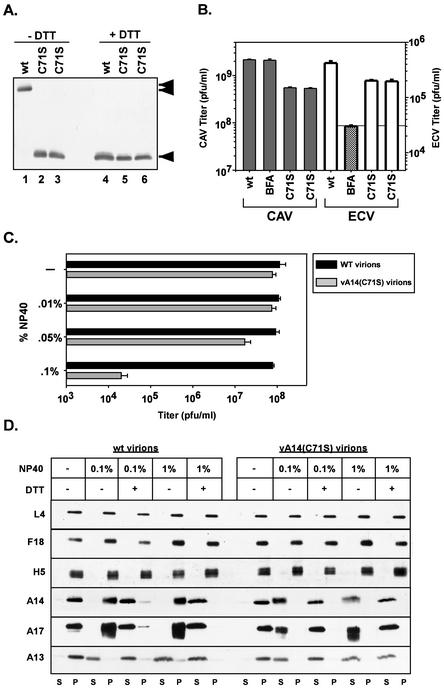
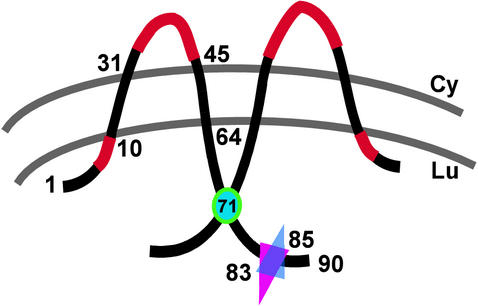
We have previously reported the construction and characterization of an inducible recombinant virus in which expression of the vaccinia virus membrane protein A14 is experimentally regulated using the tetracycline operator-repressor system. Repression of A14, which results in a 1,000-fold reduction in viral yield, leads to an early block in viral morphogenesis characterized by the accumulation of large virosomes, empty "crescents" that fail to contact these virosomes, and, most strikingly, large numbers of aberrant 25-nm vesicles. Here we report the establishment of a transient-complementation system for the structure-function analysis of A14. We have constructed numerous mutant alleles of A14 designed to identify and test the importance of key structural and sequence motifs within A14, including sites of posttranslational modification, such as glycosylation, phosphorylation, and dimerization. From these studies we have determined that robust complementation ability requires an intact N terminus and two regions flanking the first membrane-spanning domain of A14. We show that A14 is modified by N-linked glycosylation both in vitro and in vivo. However, only a minority of A14 molecules are glycosylated in vivo and these are not encapsidated. In this report we also identify the sole phosphorylated serine residue of A14 as lying within the NHS(85) motif that undergoes glycosylation. Additionally, we show that the Cys(71) residue is required for intermolecular disulfide bond formation and describe the properties of a virus expressing an allele of A14 that cannot form disulfide-linked dimers.
Figures
Similar articles
-
Elucidating the essential role of the A14 phosphoprotein in vaccinia virus morphogenesis: construction and characterization of a tetracycline-inducible recombinant.J Virol. 2000 Apr;74(8):3682-95. doi: 10.1128/jvi.74.8.3682-3695.2000. J Virol. 2000. PMID: 10729144 Free PMC article.
-
Biogenesis of the vaccinia virus membrane: genetic and ultrastructural analysis of the contributions of the A14 and A17 proteins.J Virol. 2013 Jan;87(2):1083-97. doi: 10.1128/JVI.02529-12. Epub 2012 Nov 7. J Virol. 2013. PMID: 23135725 Free PMC article.
-
Vaccinia virus morphogenesis: a13 phosphoprotein is required for assembly of mature virions.J Virol. 2004 Aug;78(16):8885-901. doi: 10.1128/JVI.78.16.8885-8901.2004. J Virol. 2004. PMID: 15280497 Free PMC article.
-
Vaccinia virus envelope H3L protein binds to cell surface heparan sulfate and is important for intracellular mature virion morphogenesis and virus infection in vitro and in vivo.J Virol. 2000 Apr;74(7):3353-65. doi: 10.1128/jvi.74.7.3353-3365.2000. J Virol. 2000. PMID: 10708453 Free PMC article.
-
The vaccinia virus A14.5L gene encodes a hydrophobic 53-amino-acid virion membrane protein that enhances virulence in mice and is conserved among vertebrate poxviruses.J Virol. 2000 May;74(9):4085-92. doi: 10.1128/jvi.74.9.4085-4092.2000. J Virol. 2000. PMID: 10756020 Free PMC article.
Cited by
-
Analysis of viral membranes formed in cells infected by a vaccinia virus L2-deletion mutant suggests their origin from the endoplasmic reticulum.J Virol. 2013 Feb;87(3):1861-71. doi: 10.1128/JVI.02779-12. Epub 2012 Nov 28. J Virol. 2013. PMID: 23192873 Free PMC article.
-
From crescent to mature virion: vaccinia virus assembly and maturation.Viruses. 2014 Oct 7;6(10):3787-808. doi: 10.3390/v6103787. Viruses. 2014. PMID: 25296112 Free PMC article. Review.
-
Mimicry Embedding Facilitates Advanced Neural Network Training for Image-Based Pathogen Detection.mSphere. 2020 Sep 9;5(5):e00836-20. doi: 10.1128/mSphere.00836-20. mSphere. 2020. PMID: 32907956 Free PMC article.
-
Vaccinia virus H7 protein contributes to the formation of crescent membrane precursors of immature virions.J Virol. 2009 Sep;83(17):8439-50. doi: 10.1128/JVI.00877-09. Epub 2009 Jun 24. J Virol. 2009. PMID: 19553304 Free PMC article.
-
Viral miniproteins.Annu Rev Microbiol. 2014;68:21-43. doi: 10.1146/annurev-micro-091313-103727. Epub 2014 Apr 10. Annu Rev Microbiol. 2014. PMID: 24742054 Free PMC article. Review.
References
-
- Bogan, A. A., and K. S. Thorn. 1998. Anatomy of hot spots in protein interfaces. J. Mol. Biol. 280:1-9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
