

Three-dimensional structure of the M-MuLV CA protein on a lipid monolayer: a general model for retroviral capsid assembly
- PMID: 12805204
- PMCID: PMC162131
- DOI: 10.1093/emboj/cdg276
Three-dimensional structure of the M-MuLV CA protein on a lipid monolayer: a general model for retroviral capsid assembly
Abstract
Although retroviruses from different genera form morphologically distinct capsids, we have proposed that all of these structures are composed of similar hexameric arrays of capsid (CA) protein subunits and that their distinct morphologies reflect different distributions of pentameric declinations that allow the structures to close. Consistent with this model, CA proteins from both HIV-1 and Rous sarcoma virus (RSV) form similar hexagonal lattices. However, recent structural studies have suggested that the Moloney murine leukemia virus (M-MuLV) CA protein may assemble differently. We now report an independent three-dimensional reconstruction of two-dimensional crystals of M-MuLV CA. This new reconstruction reveals a hexameric lattice that is similar to those formed by HIV-1 and RSV CA, supporting a generalized model for retroviral capsid assembly.
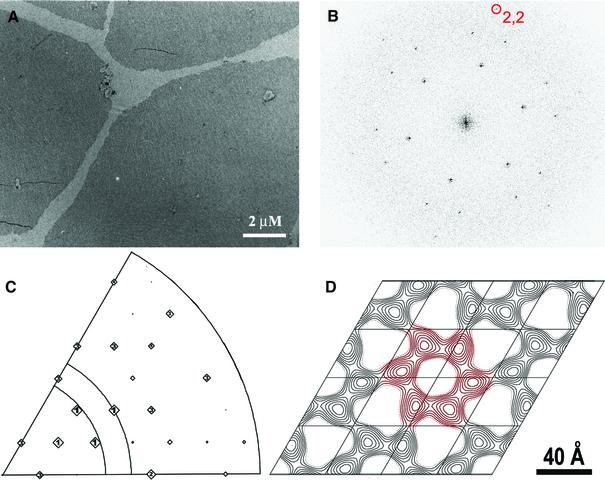
Figures



Similar articles
-
Consideration of the three-dimensional structure of core shells (capsids) in spherical retroviruses.Micron. 2007;38(5):462-70. doi: 10.1016/j.micron.2006.11.007. Epub 2006 Dec 19. Micron. 2007. PMID: 17223564
-
A small loop in the capsid protein of Moloney murine leukemia virus controls assembly of spherical cores.J Virol. 2006 Mar;80(6):2884-93. doi: 10.1128/JVI.80.6.2884-2893.2006. J Virol. 2006. PMID: 16501097 Free PMC article.
-
Retrovirus capsid protein assembly arrangements.J Mol Biol. 2003 Jan 3;325(1):225-37. doi: 10.1016/s0022-2836(02)01176-2. J Mol Biol. 2003. PMID: 12473464
-
The capsid protein of human immunodeficiency virus: intersubunit interactions during virus assembly.FEBS J. 2009 Nov;276(21):6098-109. doi: 10.1111/j.1742-4658.2009.07313.x. FEBS J. 2009. PMID: 19825044 Review.
-
Structural analyses of Phycodnaviridae and Iridoviridae.Acta Crystallogr D Biol Crystallogr. 2003 Dec;59(Pt 12):2053-9. doi: 10.1107/s090744490302225x. Epub 2003 Nov 27. Acta Crystallogr D Biol Crystallogr. 2003. PMID: 14646061 Review.
Cited by
-
Spontaneous non-canonical assembly of CcmK hexameric components from β-carboxysome shells of cyanobacteria.PLoS One. 2017 Sep 21;12(9):e0185109. doi: 10.1371/journal.pone.0185109. eCollection 2017. PLoS One. 2017. PMID: 28934279 Free PMC article.
-
The N-terminus of murine leukaemia virus p12 protein is required for mature core stability.PLoS Pathog. 2014 Oct 30;10(10):e1004474. doi: 10.1371/journal.ppat.1004474. eCollection 2014 Oct. PLoS Pathog. 2014. PMID: 25356837 Free PMC article.
-
Contribution of glutamine residues in the helix 4-5 loop to capsid-capsid interactions in simian immunodeficiency virus of macaques.J Virol. 2014 Sep;88(18):10289-302. doi: 10.1128/JVI.01388-14. Epub 2014 Jul 2. J Virol. 2014. PMID: 24991000 Free PMC article.
-
Lipid monolayer and sparse matrix screening for growing two-dimensional crystals for electron crystallography: methods and examples.Methods Mol Biol. 2013;955:527-37. doi: 10.1007/978-1-62703-176-9_28. Methods Mol Biol. 2013. PMID: 23132079 Free PMC article.
-
Unclosed HIV-1 capsids suggest a curled sheet model of assembly.J Mol Biol. 2013 Jan 9;425(1):112-23. doi: 10.1016/j.jmb.2012.10.006. Epub 2012 Oct 16. J Mol Biol. 2013. PMID: 23079241 Free PMC article.
References
-
- Barklis E., McDermott,J., Wilkens,S., Fuller,S. and Thompson,D. (1998) Organization of HIV-1 capsid proteins on a lipid monolayer. J. Biol. Chem., 273, 7177–7180. - PubMed
-
- Campos-Olivas R., Newman,J.L. and Summers,M.F. (2000) Solution structure and dynamics of the Rous sarcoma virus capsid protein and comparison with capsid proteins of other retroviruses. J. Mol. Biol., 296, 633–649. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
