

The Drosophila hairy RNA localization signal modulates the kinetics of cytoplasmic mRNA transport
- PMID: 12743042
- PMCID: PMC155988
- DOI: 10.1093/emboj/cdg230
The Drosophila hairy RNA localization signal modulates the kinetics of cytoplasmic mRNA transport
Abstract
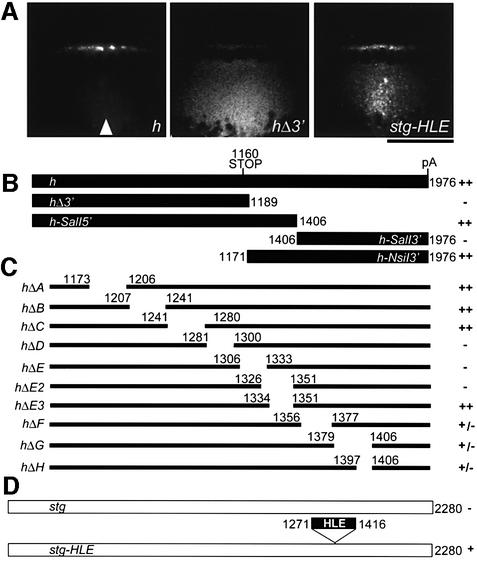
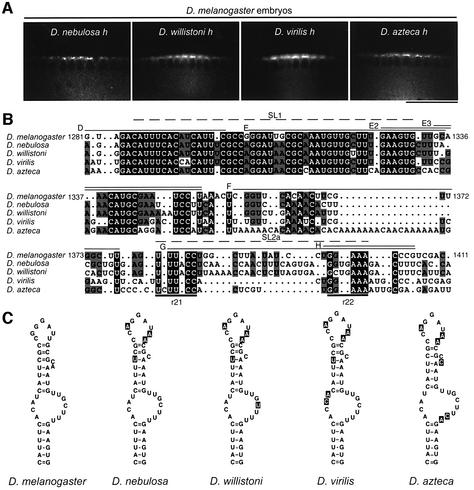
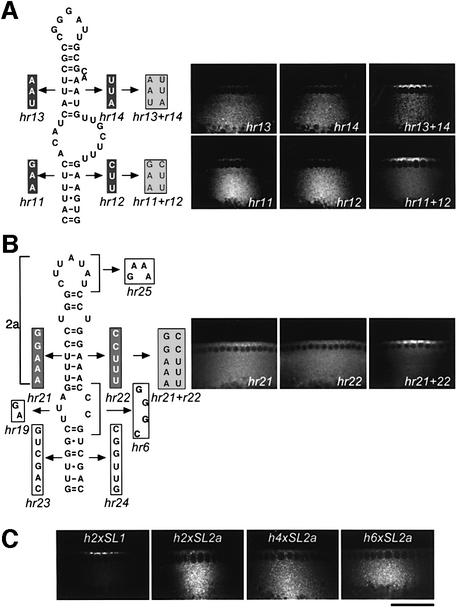
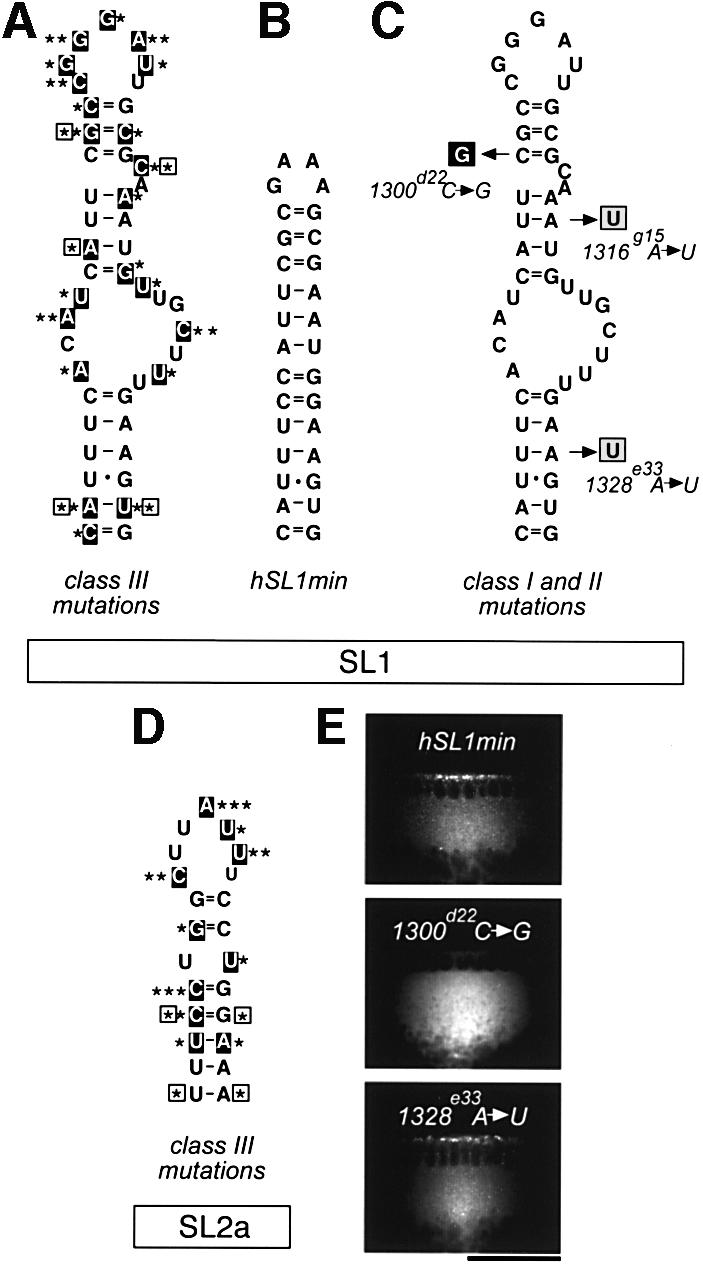
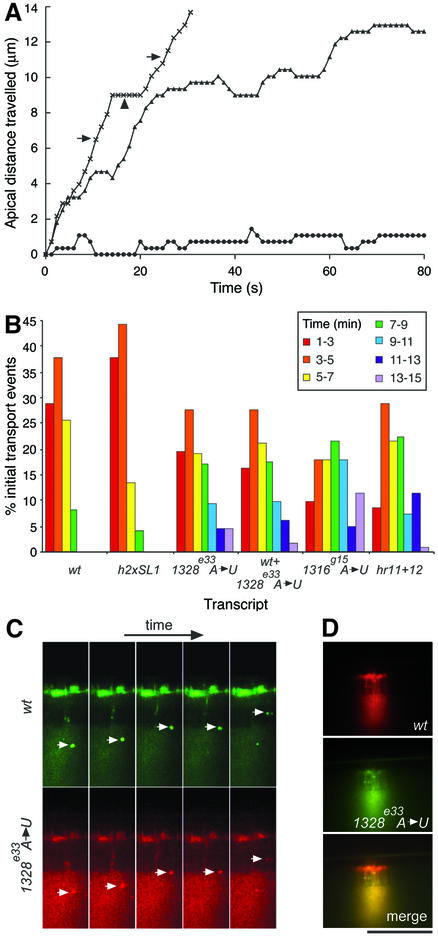
In several Drosophila cell types, mRNA transport depends on microtubules, the molecular motor dynein and trans-acting factors including Egalitarian and Bicaudal-D. However, the molecular basis of transcript recognition by the localization machinery is poorly understood. Here, we characterize the features of hairy pair-rule RNA transcripts that mediate their apical localization, using in vivo injection of fluorescently labelled mRNAs into syncytial blastoderm embryos. We show that a 121-nucleotide element within the 3'-untranslated region is necessary and sufficient to mediate apical transport. The signal comprises two essential stem-loop structures, in which double-stranded stems are crucial for localization. Base-pair identities within the stems are not essential, but can contribute to the efficiency of localization, suggesting that specificity is mediated by higher-order structure. Using time-lapse microscopy, we measure the kinetics of localization and show that impaired localization of mutant signals is due to delayed formation of active motor complexes and, unexpectedly, to slower movement. These findings, and those from co-injecting wild-type and mutant RNAs, suggest that the efficiency of molecular motors is modulated by the character of their cargoes.
Figures

Similar articles
-
Conserved signals and machinery for RNA transport in Drosophila oogenesis and embryogenesis.Nature. 2001 Dec 6;414(6864):611-6. doi: 10.1038/414611a. Nature. 2001. PMID: 11740552
-
A stem-loop structure in the wingless transcript defines a consensus motif for apical RNA transport.Development. 2008 Jan;135(1):133-43. doi: 10.1242/dev.014068. Epub 2007 Nov 28. Development. 2008. PMID: 18045835
-
Clathrin heavy chain plays multiple roles in polarizing the Drosophila oocyte downstream of Bic-D.Development. 2014 May;141(9):1915-26. doi: 10.1242/dev.099432. Epub 2014 Apr 9. Development. 2014. PMID: 24718986
-
mRNA localization in metazoans: A structural perspective.RNA Biol. 2017 Nov 2;14(11):1473-1484. doi: 10.1080/15476286.2017.1338231. Epub 2017 Jul 31. RNA Biol. 2017. PMID: 28640665 Free PMC article. Review.
-
Establishing and maintaining cell polarity with mRNA localization in Drosophila.Bioessays. 2016 Mar;38(3):244-53. doi: 10.1002/bies.201500088. Epub 2016 Jan 15. Bioessays. 2016. PMID: 26773560 Free PMC article. Review.
Cited by
-
RNA:RNA interaction can enhance RNA localization in Drosophila oocytes.RNA. 2012 Apr;18(4):729-37. doi: 10.1261/rna.026674.111. Epub 2012 Feb 17. RNA. 2012. PMID: 22345148 Free PMC article.
-
The dynamic duo of microtubule polymerase Mini spindles/XMAP215 and cytoplasmic dynein is essential for maintaining Drosophila oocyte fate.Proc Natl Acad Sci U S A. 2023 Sep 26;120(39):e2303376120. doi: 10.1073/pnas.2303376120. Epub 2023 Sep 18. Proc Natl Acad Sci U S A. 2023. PMID: 37722034 Free PMC article.
-
A stem-loop structure directs oskar mRNA to microtubule minus ends.RNA. 2014 Apr;20(4):429-39. doi: 10.1261/rna.041566.113. Epub 2014 Feb 26. RNA. 2014. PMID: 24572808 Free PMC article.
-
Identifying and searching for conserved RNA localisation signals.Methods Mol Biol. 2011;714:447-66. doi: 10.1007/978-1-61779-005-8_27. Methods Mol Biol. 2011. PMID: 21431757 Free PMC article.
-
Staufen targets coracle mRNA to Drosophila neuromuscular junctions and regulates GluRIIA synaptic accumulation and bouton number.Dev Biol. 2014 Aug 15;392(2):153-67. doi: 10.1016/j.ydbio.2014.06.007. Epub 2014 Jun 19. Dev Biol. 2014. PMID: 24951879 Free PMC article.
References
-
- Beverley S.M. and Wilson,A.C. (1984) Molecular evolution in Drosophila and the higher Diptera II. A time scale for fly evolution. J. Mol. Evol., 21, 1–13. - PubMed
-
- Bullock S.L. and Ish-Horowicz,D. (2001) Conserved signals and machinery for RNA transport in Drosophila oogenesis and embryogenesis. Nature, 414, 611–616. - PubMed
-
- Chartrand P., Meng,X.H., Singer,R.H. and Long,R.M. (1999) Structural elements required for the localization of ASH1 mRNA and of a green fluorescent protein reporter particle in vivo. Curr. Biol., 9, 333–336. - PubMed
-
- Davis I. and Ish-Horowicz,D. (1991) Apical localization of pair-rule transcripts requires 3′ sequences and limits protein diffusion in the Drosophila blastoderm embryo. Cell, 67, 927–940. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
