

Rapid invasion of host cells by Toxoplasma requires secretion of the MIC2-M2AP adhesive protein complex
- PMID: 12727875
- PMCID: PMC156089
- DOI: 10.1093/emboj/cdg217
Rapid invasion of host cells by Toxoplasma requires secretion of the MIC2-M2AP adhesive protein complex
Abstract
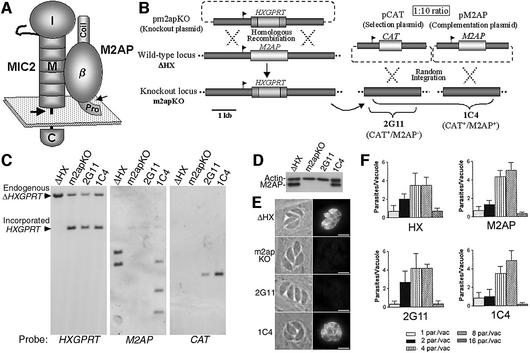
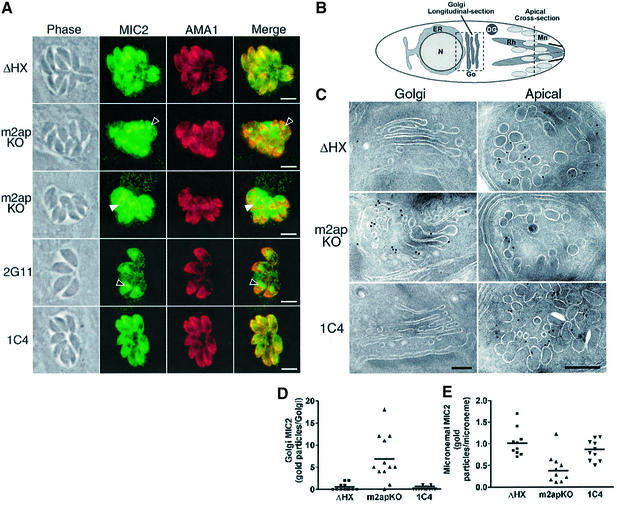
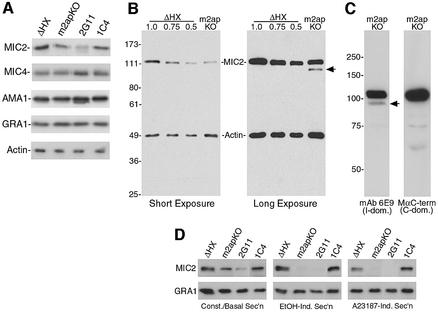
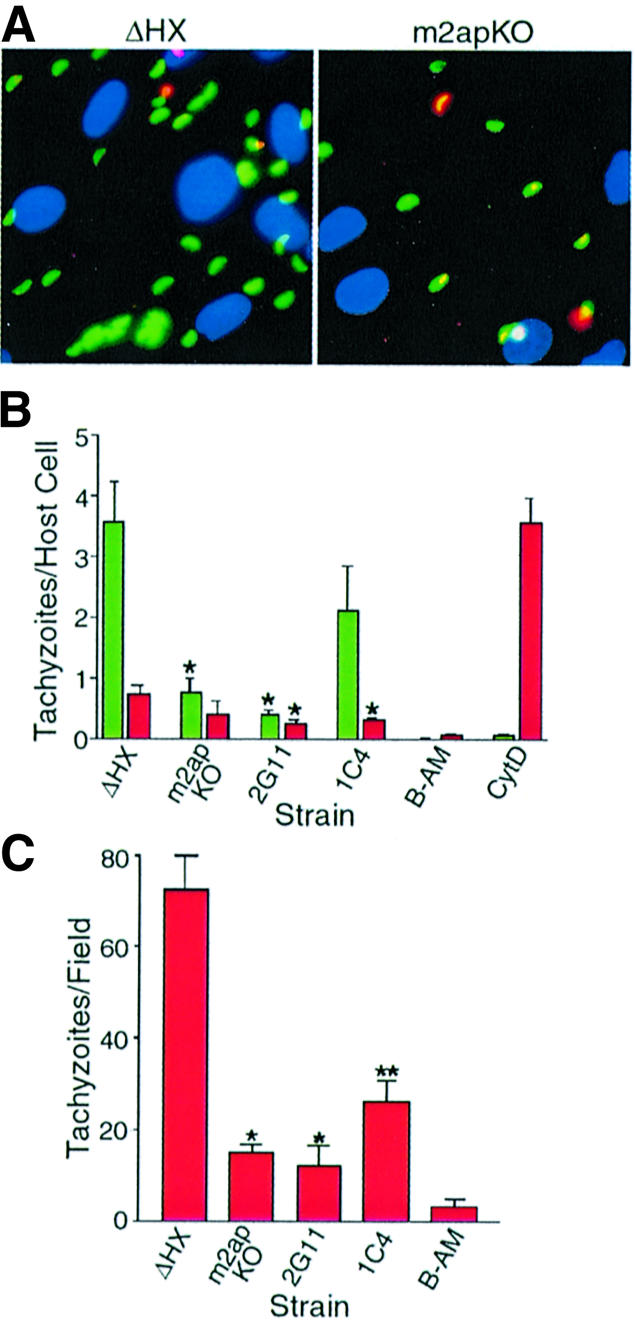
Vertebrate cells are highly susceptible to infection by obligate intracellular parasites such as Toxoplasma gondii, yet the mechanism by which these microbes breach the confines of their target cell is poorly understood. While it is thought that Toxoplasma actively invades by secreting adhesive proteins from internal organelles called micronemes, no genetic evidence is available to support this contention. Here, we report successful disruption of M2AP, a microneme protein tightly associated with an adhesive protein called MIC2. M2AP knockout parasites were >80% impaired in host cell entry. This invasion defect was likely due to defective expression of MIC2, which partially accumulated in the parasite endoplasmic reticulum and Golgi. M2AP knockout parasites were also unable to rapidly secrete MIC2, an event that normally accompanies parasite attachment to a target cell. These findings indicate a critical role for the MIC2-M2AP protein complex in parasite invasion.
Figures
Similar articles
-
Preparing for an invasion: charting the pathway of adhesion proteins to Toxoplasma micronemes.Parasitol Res. 2006 Apr;98(5):389-95. doi: 10.1007/s00436-005-0062-2. Epub 2005 Dec 30. Parasitol Res. 2006. PMID: 16385407 Review.
-
Toxoplasma MIC2 is a major determinant of invasion and virulence.PLoS Pathog. 2006 Aug;2(8):e84. doi: 10.1371/journal.ppat.0020084. PLoS Pathog. 2006. PMID: 16933991 Free PMC article.
-
The toxoplasma proteins MIC2 and M2AP form a hexameric complex necessary for intracellular survival.J Biol Chem. 2004 Mar 5;279(10):9362-9. doi: 10.1074/jbc.M312590200. Epub 2003 Dec 10. J Biol Chem. 2004. PMID: 14670959
-
A cleavable propeptide influences Toxoplasma infection by facilitating the trafficking and secretion of the TgMIC2-M2AP invasion complex.Mol Biol Cell. 2006 Oct;17(10):4551-63. doi: 10.1091/mbc.e06-01-0064. Epub 2006 Aug 16. Mol Biol Cell. 2006. PMID: 16914527 Free PMC article.
-
Toxoplasma gondii: microneme protein MIC2.Int J Biochem Cell Biol. 2005 Nov;37(11):2266-72. doi: 10.1016/j.biocel.2005.06.006. Int J Biochem Cell Biol. 2005. PMID: 16084754 Review.
Cited by
-
Host E3 ubiquitin ligase ITCH mediates Toxoplasma gondii effector GRA35-triggered NLRP1 inflammasome activation and cell-autonomous immunity.bioRxiv [Preprint]. 2023 Dec 14:2023.12.13.571530. doi: 10.1101/2023.12.13.571530. bioRxiv. 2023. Update in: mBio. 2024 Mar 13;15(3):e0330223. doi: 10.1128/mbio.03302-23 PMID: 38168400 Free PMC article. Updated. Preprint.
-
Fetuin-A, a hepatocyte-specific protein that binds Plasmodium berghei thrombospondin-related adhesive protein: a potential role in infectivity.Infect Immun. 2005 Sep;73(9):5883-91. doi: 10.1128/IAI.73.9.5883-5891.2005. Infect Immun. 2005. PMID: 16113307 Free PMC article.
-
GRA proteins of Toxoplasma gondii: maintenance of host-parasite interactions across the parasitophorous vacuolar membrane.Korean J Parasitol. 2009 Oct;47 Suppl(Suppl):S29-37. doi: 10.3347/kjp.2009.47.S.S29. Korean J Parasitol. 2009. PMID: 19885333 Free PMC article. Review.
-
Thrombospondin Related Anonymous Protein Superfamily in Vector-Borne Apicomplexans: The Parasite's Toolkit for Cell Invasion.Front Cell Infect Microbiol. 2022 Apr 6;12:831592. doi: 10.3389/fcimb.2022.831592. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35463644 Free PMC article. Review.
-
TgMORN1 is a key organizer for the basal complex of Toxoplasma gondii.PLoS Pathog. 2010 Feb 5;6(2):e1000754. doi: 10.1371/journal.ppat.1000754. PLoS Pathog. 2010. PMID: 20140195 Free PMC article.
References
-
- Arvan P., Zhao,X., Ramos-Castaneda,J. and Chang,A. (2002) Secretory pathway quality control operating in Golgi, plasmalemmal, and endosomal systems. Traffic, 3, 771–780. - PubMed
-
- Blond-Elguindi S., Cwirla,S.E., Dower,W.J., Lipshutz,R.J., Sprang,S.R., Sambrook,J.F. and Gething,M.J. (1993) Affinity panning of a library of peptides displayed on bacteriophages reveals the binding specificity of BiP. Cell, 75, 717–728. - PubMed
-
- Brecht S., Carruthers,V.B., Ferguson,D.J., Giddings,O.K., Wang,G., Jaekle,U., Harper,J.M., Sibley,L.D. and Soldati,D. (2001) The Toxoplasma micronemal protein MIC4 is an adhesin composed of six conserved apple domains. J. Biol. Chem., 276, 4119–4127. - PubMed
-
- Brossier J., Jewett,T.J., Lovett,J.L. and Sibley,L.D. (2003) C-terminal processing of the Toxoplasma protein MIC2 is essential for invasion of host cells. J. Biol. Chem., 278, 6229–6234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
