

Review
doi: 10.1038/nature01602.
Adaptation of core mechanisms to generate cell polarity
Affiliations
- PMID: 12700771
- PMCID: PMC3373010
- DOI: 10.1038/nature01602
Item in Clipboard
Review
Adaptation of core mechanisms to generate cell polarity
Nature.
.
Abstract
Cell polarity is defined as asymmetry in cell shape, protein distributions and cell functions. It is characteristic of single-cell organisms, including yeast and bacteria, and cells in tissues of multi-cell organisms such as epithelia in worms, flies and mammals. This diversity raises several questions: do different cell types use different mechanisms to generate polarity, how is polarity signalled, how do cells react to that signal, and how is structural polarity translated into specialized functions? Analysis of evolutionarily diverse cell types reveals that cell-surface landmarks adapt core pathways for cytoskeleton assembly and protein transport to generate cell polarity.
Figures
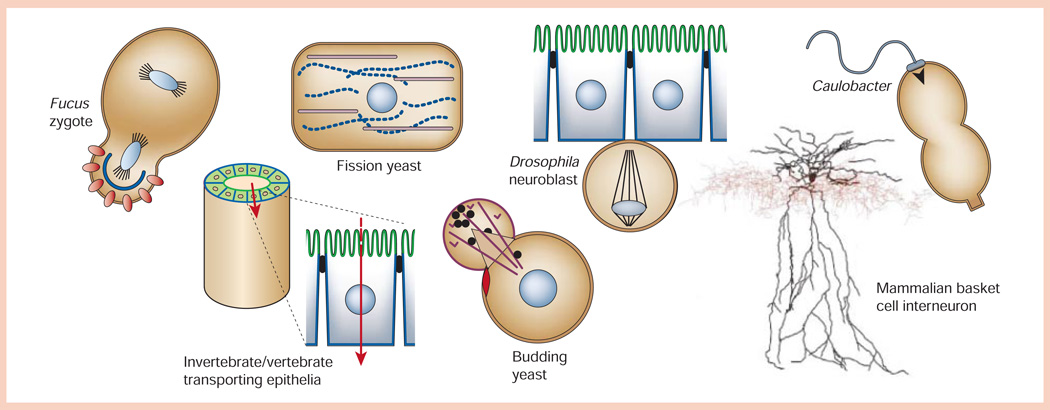
Diversity of shapes of polarized cells (not to scale). Fucus zygote exposed to a light gradient showing polarized distributions of ion channels/dihydropyridine receptors (red circles) and F-actin (blue line) in the rhizoid cell (bottom) compared to the thallus cell (top). Fission yeast (Schizosaccharomyces pombe) showing polarized distributions of actin (purple) and microtubules (blue dotted line) in the long axis of the cell, and the nucleus positioned in the centre of the cell. Drosophila neuroblast delaminated from the ventral neuroectoderm with an asymmetric plane of division that will yield a large ‘apical’ neuroblast stem cell and a small ‘basal’ ganglion mother cell. Caulobacter crescentus predivisional cell showing polarized distributions of the flagellum (swarmer cell, top) and stalk (stalked cell, bottom). Invertebrate/vertebrate transporting epithelium showing organization of polarized epithelial cells (apical membrane, green; basolateral membrane, blue) in a tube that separates two biological compartments and regulates vectorial transport of ions/solutes (red arrow) between those compartments. Budding yeast (Saccharomyces cerevisiae) forming a daughter cell ‘bud’ from the mother cell next to the previous site of cytokinesis (bud scar, red disc), and orienting actin cables (purple) for transport of vesicles (black circles) from the mother to daughter cell. Mammalian basket cell interneuron showing the distribution of the soma/dendrite (black) and axon (red; image courtesy of D. Madison, Stanford University School of Medicine).
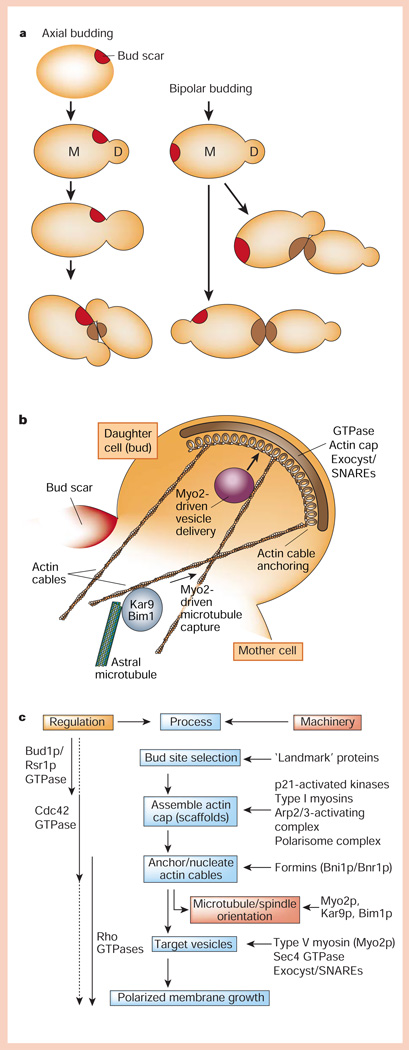
Protein pathways for generating cell polarity in budding yeast. a, Axial and bipolar budding patterns. b, A complex of proteins is assembled at the bud tip that orients the actin cytoskeleton, astral microtubules and vesicle delivery to the bud. c, Hierarchical organization of regulators, processes and cellular machinery linking bud-site selection to assembly of an actin cap, anchoring/nucleation of actin cables, and vesicle targeting that results in polarized membrane growth at the bud.
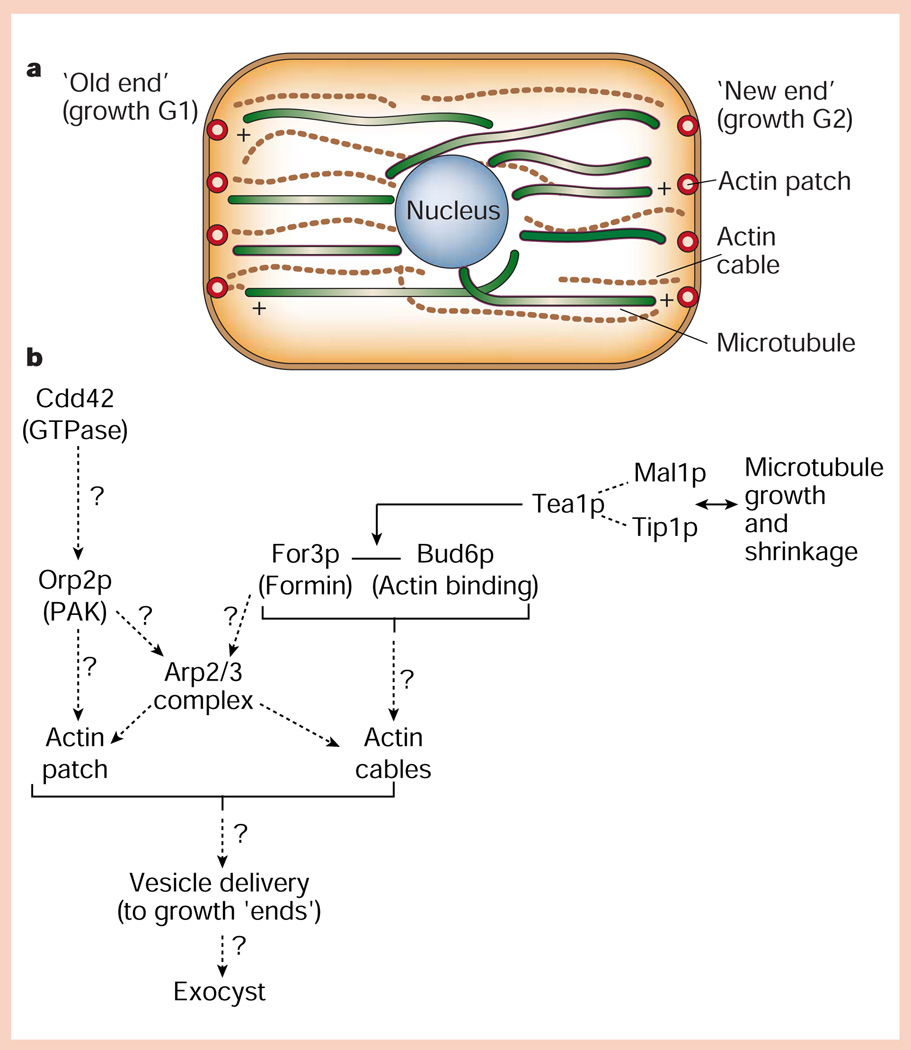
Protein pathways for generating cell polarity in fission yeast. a, Distribution of actin (cables and cortical patches) and microtubule cytoskeletons relative to the ‘old’ and ‘new’ ends of the cell, which grow during the G1 and G2 phases of the cell cycle, respectively. b, Hypothetical organization of proteins in cortical actin patches, and interactions of microtubule plus ends with the cell end/cortical actin patch (for details, see text).
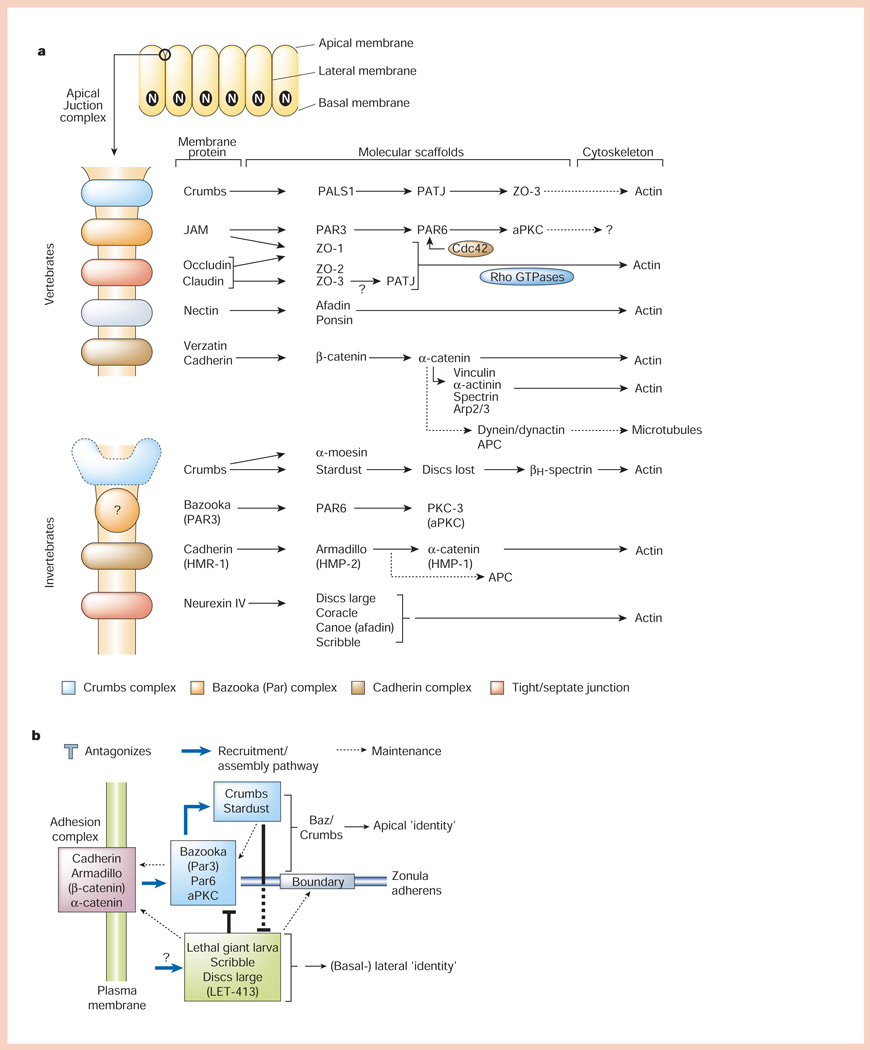
Organization of polarized epithelial cells and the apical junctional complex. a, Polarized epithelial cells form a monolayer in which the apical (unbounded surface) is separated at the boundary with the basal and lateral membranes (bounded surfaces) by the apical junctional complex (top). The main part of the panel shows molecular organization of the apical junctional complex. In vertebrates, the apical junctional complex is separated into structurally and functionally different sub-domains comprising membrane proteins (Crumbs, JAM (junctional adhesion molecule), nectin, occludin/claudin and cadherin) linked to modular protein scaffolds, which in turn bind mostly to the actin cytoskeleton, although links to microtubules are possible. In invertebrates (C. elegans and Drosophila), the apical junctional complex is similarly organized, except that the ‘tight junction’ function is provided by the septate junction localized below the cadherin (adherens) junction. b, Simplified scheme for how different protein complexes in the apical junctional complex regulate cell–cell adhesion (cadherin complex), and apical membrane (Bazooka and Crumbs complexes) and lateral membrane (Lethal giant larvae, Scribble and Disc large complex) identity. For details see text.
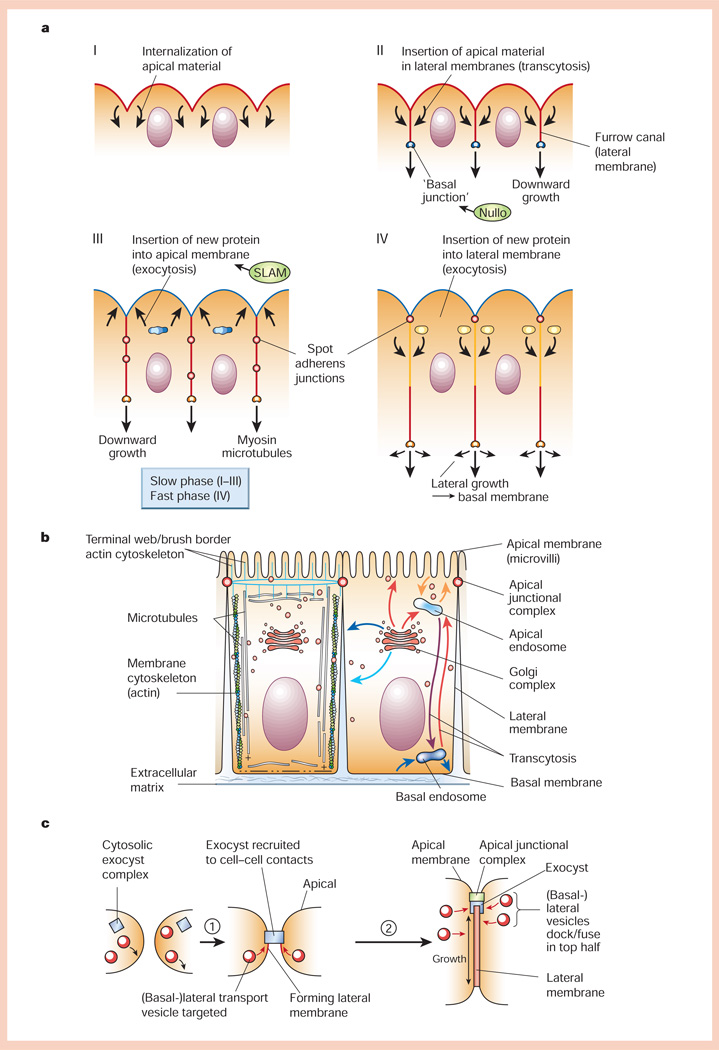
Generation of cell polarity in epithelia. a, Formation of the cellular blastoderm in early Drosophila embryogenesis (for details, see text). b, Schematic representation of polarized epithelial cells. Left, organization of the actin and microtubule cytoskeletons; right, organization of vesicle transport pathways to different plasma membrane domains either directly from the Golgi complex, or indirectly via apical or basal endosomes through endocytic or transcytotic pathways. c, Generation of the lateral membrane domain in cultured epithelial cells. Prior to cadherin-mediated cell–cell adhesion, the exocyst is cytosolic and vesicles fuse with the basal membrane. Upon cell–cell adhesion (step 1), the exocyst is recruited to cell–cell contacts and vesicles fuse with the forming lateral membrane. As more vesicles fuse, the lateral membrane increases in area around sixfold. Later (step 2), the exocyst and vesicle delivery are located in the apical region of the lateral membrane.
Comment in
-
The cytoskeleton, cellular motility and the reductionist agenda.Nature. 2003 Apr 17;422(6933):741-5. doi: 10.1038/nature01598. Nature. 2003. PMID: 12700767
Similar articles
-
Regulation of epithelial cell shape and polarity by cell-cell adhesion (Review).Mol Membr Biol. 2002 Apr-Jun;19(2):113-20. doi: 10.1080/09687680210137219. Mol Membr Biol. 2002. PMID: 12126229 Review.
-
Structural and regulatory functions of keratins.Exp Cell Res. 2007 Jun 10;313(10):2021-32. doi: 10.1016/j.yexcr.2007.03.005. Epub 2007 Mar 15. Exp Cell Res. 2007. PMID: 17434482 Review.
-
Dynamics and regulation of epithelial adherens junctions: recent discoveries and controversies.Int Rev Cell Mol Biol. 2013;303:27-99. doi: 10.1016/B978-0-12-407697-6.00002-7. Int Rev Cell Mol Biol. 2013. PMID: 23445808 Review.
-
Chapter 3: acquisition of membrane polarity in epithelial tube formation patterns, signaling pathways, molecular mechanisms, and disease.Int Rev Cell Mol Biol. 2009;274:129-82. doi: 10.1016/S1937-6448(08)02003-0. Int Rev Cell Mol Biol. 2009. PMID: 19349037 Review.
-
Mammalian PAR-1 determines epithelial lumen polarity by organizing the microtubule cytoskeleton.J Cell Biol. 2004 Mar 1;164(5):717-27. doi: 10.1083/jcb.200308104. Epub 2004 Feb 23. J Cell Biol. 2004. PMID: 14981097 Free PMC article.
Cited by
-
Prion Protein at the Leading Edge: Its Role in Cell Motility.Int J Mol Sci. 2020 Sep 12;21(18):6677. doi: 10.3390/ijms21186677. Int J Mol Sci. 2020. PMID: 32932634 Free PMC article. Review.
-
The plasma membrane potential and the organization of the actin cytoskeleton of epithelial cells.Int J Cell Biol. 2012;2012:121424. doi: 10.1155/2012/121424. Epub 2012 Jan 23. Int J Cell Biol. 2012. PMID: 22315611 Free PMC article.
-
MARCKS modulates radial progenitor placement, proliferation and organization in the developing cerebral cortex.Development. 2009 Sep;136(17):2965-75. doi: 10.1242/dev.036616. Development. 2009. PMID: 19666823 Free PMC article.
-
Cell polarity signalling at the birth of multicellularity: What can we learn from the first animals.Front Cell Dev Biol. 2022 Nov 24;10:1024489. doi: 10.3389/fcell.2022.1024489. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36506100 Free PMC article. Review.
-
The STE20/germinal center kinase POD6 interacts with the NDR kinase COT1 and is involved in polar tip extension in Neurospora crassa.Mol Biol Cell. 2006 Sep;17(9):4080-92. doi: 10.1091/mbc.e06-01-0072. Epub 2006 Jul 5. Mol Biol Cell. 2006. PMID: 16822837 Free PMC article.
References
-
- Chant J. Cell polarity in yeast. Annu. Rev. Cell Dev. Biol. 1999;15:365–391. - PubMed
-
- Chant J, Herskowitz I. Genetic control of bud site selection in yeast by a set of gene products that constitute a morphogenetic pathway. Cell. 1991;65:1203–1212. - PubMed
-
- Pruyne D, Bretscher A. Polarization of cell growth in yeast I. Establishment and maintenance of polarity states. J. Cell Sci. 2000;113:365–375. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
